آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| DNase Sensitivity Detects Changes in Chromatin Structure |


|
|
Read More
Date: 9-5-2016
Date: 30-11-2015
Date: 5-5-2021
|
DNase Sensitivity Detects Changes in Chromatin Structure
KEY CONCEPTS
-Hypersensitive sites are found at the promoters of expressed genes as well as other important sites such as origins of replication and centromeres.
-Hypersensitive sites are generated by the binding of factors that exclude histone octamers.
- A domain containing a transcribed gene is defined by increased sensitivity to degradation by DNase I.
Numerous changes occur to chromatin in active or potentially active regions. These include distinctive structural changes that occur at specific sites associated with initiation of transcription or with certain structural features in DNA. These changes were first detected by the effects of digestion with very low concentrations of the enzyme DNase I.
When chromatin is digested with DNase I, the first effect is the introduction of breaks in the duplex at specific, hypersensitive sites. Susceptibility to DNase I reflects the availability of DNA in chromatin; thus, these sites represent chromatin regions in which the DNA is particularly exposed because it is not organized in the usual nucleosomal structure. A typical hypersensitive site is 100 times more sensitive to enzyme attack than bulk chromatin. These sites are also hypersensitive to other nucleases and to chemical agents.
Hypersensitive sites are created by the local structure of chromatin, which can be tissue specific. Researchers can determine their locations by the technique of indirect end labeling that we introduced earlier in the context of nucleosome positioning. This application of the technique is recapitulated in FIGURE 1.
In this case, cleavage at the hypersensitive site by DNase I is used to generate one end of the fragment. Its distance is measured from the other end, which is generated by cleavage with a restriction enzyme.
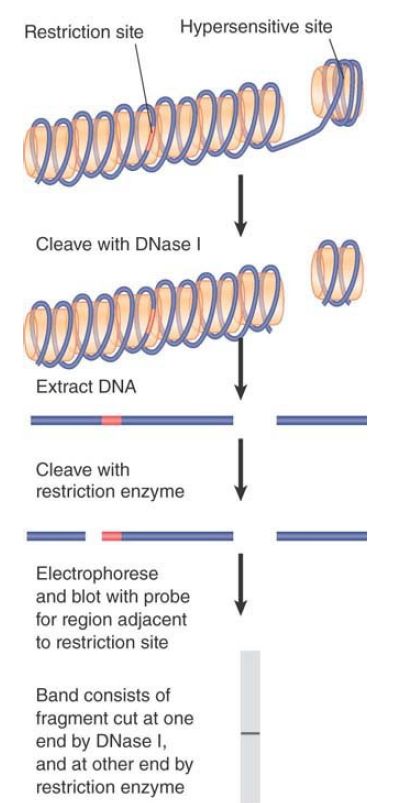
FIGURE 1. Indirect end labeling identifies the distance of a DNase hypersensitive site from a restriction cleavage site. The existence of a particular cutting site for DNase I generates a discrete fragment, whose size indicates the distance of the DNase I hypersensitive site from the restriction site.
Many hypersensitive sites are related to gene expression. Every active gene has a hypersensitive site, or sometimes more than one, in the region of the promoter. Most hypersensitive sites are found only in chromatin of cells in which the associated gene is either being expressed or is poised for expression; they do not occur when the gene is inactive. The 5′ hypersensitive site(s) appear before transcription begins and occur in DNA sequences that are required for gene expression.
What is the structure of a hypersensitive site? Its preferential accessibility to nucleases indicates that it is not protected by histone octamers, but this does not necessarily imply that it is free of protein. A region of free DNA might be vulnerable to damage, and would be unable to exclude nucleosomes. In fact, hypersensitive sites typically result from the binding of specific regulatory proteins that exclude nucleosomes. It is very common to find pairs of hypersensitive sites that flank a nuclease-resistant core; the binding of nucleosome-excluding proteins is probably the basis for the existence of the protected region within the hypersensitive sites.
The proteins that generate hypersensitive sites are likely to be regulatory factors of various types, because hypersensitive sites are found associated with promoters and other elements that regulate transcription, origins of replication, centromeres, and sites with other structural significance. In some cases, they are associated with more extensive organization of chromatin structure.
A hypersensitive site can provide a boundary for a series of positioned nucleosomes. Hypersensitive sites associated with transcription may be generated by transcription factors when they bind to the promoter as part of the process that makes it accessible to RNA polymerase.
In addition to detecting hypersensitive sites, researchers also can use DNase I digestion to assess the relative accessibility of a genomic region. A region of the genome that contains an active gene can have an altered overall structure, often typified by a general increase in overall DNase sensitivity, in addition to specific hypersensitive sites. The change in structure precedes, and is different from, the disruption of nucleosome structure that might be caused by the actual passage of RNA polymerase. DNase I sensitivity defines a chromosomal domain, which is a region of altered structure including at least one active transcription unit, and sometimes extending farther. (Note that use of the term domain does not imply any necessary connection with the structural domains identified by the loops of chromatin or chromosomes.)
When chromatin is extensively digested with DNase I, it is eventually degraded into very small fragments of DNA. The fate of individual genes can be followed by quantitating the amount of DNA that survives to react with a specific probe. The protocol is outlined in FIGURE 2. The principle is that the loss of a particular band indicates that the corresponding region of DNA has been degraded by the enzyme.
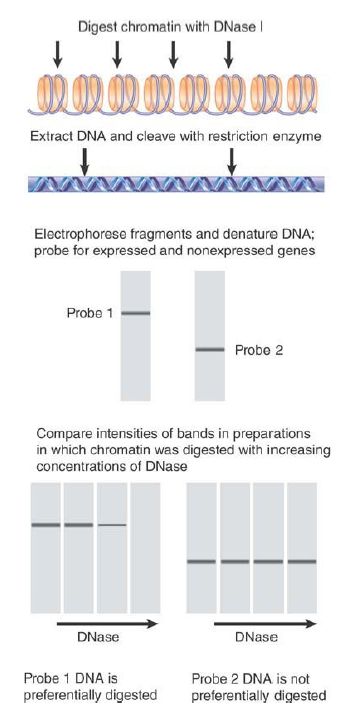
FIGURE 2. Sensitivity to DNase I can be measured by determining the rate of disappearance of the material hybridizing with a particular probe.
Studies using these methods reveal that the bulk of chromatin is relatively resistant to DNase I and contains nonexpressed genes (as well as other sequences). A gene becomes relatively susceptible to nuclease digestion specifically in the tissue(s) in which it is expressed or is poised to be expressed, and remains nuclease resistant in lineages in which the gene is silent.
What is the extent of a preferentially sensitive region? Researchers can determine this by using a series of probes representing the flanking regions and the transcription unit itself. The sensitive region always extends over the entire transcribed region; an additional region of several kb on either side might show an intermediate level of sensitivity (probably as the result of spreading effects).
The critical concept implicit in the description of the domain is that a region of high sensitivity to DNase I extends over a considerable distance. Often we think of regulation as residing in events that occur at a discrete site in DNA—for example, in the ability to initiate transcription at the promoter. Even if this is true, such regulation must determine, or must be accompanied by, a more wide-ranging change in structure.



|
|
|
|
إجراء أول اختبار لدواء "ثوري" يتصدى لعدة أنواع من السرطان
|
|
|
|
|
|
|
دراسة تكشف "سببا غريبا" يعيق نمو الطيور
|
|
|
|
|
|
لأعضاء مدوّنة الكفيل السيد الصافي يؤكّد على تفعيل القصة والرواية المجسّدة للمبادئ الإسلامية والموجدة لحلول المشاكل المجتمعية
|
|
|
|
قسم الشؤون الفكرية يناقش سبل تعزيز التعاون المشترك مع المؤسّسات الأكاديمية في نيجيريا
|
|
|
|
ضمن برنامج عُرفاء المنصّة قسم التطوير يقيم ورشة في (فنّ الٕالقاء) لمنتسبي العتبة العباسية
|
|
|
|
وفد نيجيري يُشيد بمشروع المجمع العلمي لحفظ القرآن الكريم
|