
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Fibrinogen |


|
|
Read More
Date: 20-5-2021
Date: 16-5-2016
Date: 10-5-2016
|
Fibrinogen
Blood clots when soluble fibrinogen is cleaved by thrombin to generate fibrin monomers which polymerize spontaneously, non-covalently and rapidly to form the insoluble fibrin clot, which is further stabilized by covalent cross-linking catalyzed by the transglutaminase, factor XIIIa . This remarkable property is dependent on the symmetrical multi-chain structure of fibrinogen, the proteolytic exposure of cryptic binding sites, and the high concentration of fibrinogen in blood plasma (2–4 mg/mL). The cessation of bleeding is further enhanced by fibrin acting as a cofactor in blood platelet aggregation. The fibrin clot further acts as a provisional matrix for the subsequent repair of the wound.
Fibrinogen is a disulfide-bonded hexameric protein, arranged as a dimer of Aa, Bb and g chains. These are the products of three distinct, but related, genes located in close association on human chromosome 4. The mature chains are 625, 461 and 411 residues, respectively, resulting in a molecular weight of 350 kDa for the hexamer. The gross topological structures of fibrinogen and fibrin have been deduced by both biochemical and biophysical methods. The overall appearance of fibrinogen is a linear trinodular structure 475 Å in length, with each nodule separated by an intervening rod-like region (1) (Figure 1). The N-terminal regions of all six chains are contained in the highly disulfide-bonded central nodule, the N-terminal disulfide knot. The N-termini of the Aa and Bb chains protrude from the nodule, while the g chains have shorter N-terminal sequences. Triple-chained coiled coils connect the central to the distal nodules, each of which is composed of two homologous globular domains comprising the C-terminal regions of the Bb and g chains. The longer Aa chain folds back along the axis of the molecule, to end in a globular domain that interacts with the N-terminal disulfide knot.
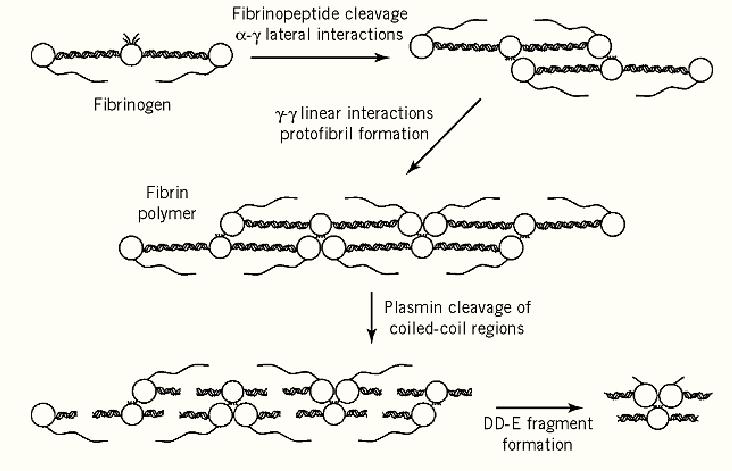
Figure 1. Schematic model of fibrin polymerization and its subsequent dissolution.
Structures have been determined by X-ray crystallography for various C-terminal proteolytic fragments of human fibrinogen (2, 3) and, although intact human fibrinogen has yet to be crystallized, the structure of native chicken fibrinogen has been resolved at a resolution of 2.7 Å (4. (The triple-chained connecting regions are susceptible to proteolysis, generating two major fragments: fragment E corresponding to the central N-terminal nodule (approximately 15% of the molecule) and fragment D containing the distal globular domains (the two identical fragments accounting for approximately 50% of the molecule). Both of these fragments contain substantial portions of the cleaved coiled coil regions with the remainder of the protein accounted for by the C-terminal regions of the Aa chains.
Fibrin polymerization essentially occurs in three sequential steps. Firstly, thrombin cleaves the N-terminal regions of the Aa and Bb chains at Arg16-Gly17 and Arg14-Gly15, respectively, releasing fibrinopeptides A and B. The N-terminal sequence of the fibrin a chain (Gly17-Pro-Arg-Val) is the major determinant of fibrin polymerization which proceeds efficiently without further modification to the fibrin(ogen) structure, and synthetic peptides corresponding to this sequence can inhibit the polymerization of fibrin monomers (5). By contrast the b-chain sequence (Gly15-His-Arg-Pro, ( which is generated more slowly, although binding to fibrinogen and fragment D, cannot inhibit polymerization. Secondly, the new N-termini of fibrin monomer spontaneously interact with binding sites in the two homologous distal globular domains, with the g-chain site particularly implicated (3). Both sites are large cavities that bind a-chain Gly-Pro-Arg ligands, but the g-chain cavity is the more specific and does not bind b-chain Gly-His-Arg ligands. It is also the more distal of the two C-terminal domains and is able to intermolecularly associate in a specific “end-to-end” manner (2). Thus adjacent g-chain sites are closely situated for optimal binding of the dimeric a-chain N-terminal residues and the efficiency of polymerization derives from both a-g lateral and g-g linear interactions. The symmetry of fibrin(ogen) allows oligomerization to proceed in both directions, generating a two-molecule thick protofibril with a half-staggered overlap (Figure 1). Finally, the fibrin fibrils are reinforced by the transglutaminase activity of factor XIIIa. The g-g linear interactions are the major sites for the introduction of intermolecular g-glutamyl- -amino-lysine isopeptide crosslinks, subsequently the globular C-terminal domains of the a chains of adjacent laterally aligned protofibrils are also crosslinked, resulting in the mature, covalently stabilized fibrin mesh. Under physiological conditions these fibers are of the order of 1 µM in diameter and thus several hundred fibrin units wide.
The assembly of fibrin also reveals epitopes that are involved in its subsequent dissolution by plasmin. Conformational changes in the vicinity of the C-terminal globular domains are thought to reveal the binding sites for tPA that are responsible for the fibrin-specific potentiation of its activity. These sites are g312-324 in the C-terminal globular domain and Aa148-160, a region interacting with the Bb-chain C-terminal globular domain. The initial binding sites for plasminogen also reside in fragment D, but subsequent plasmin degradation of fibrin reveals C-terminal Lys residues that act as supplementary sites for plasminogen binding, thereby propagating plasminogen activation. Once generated plasmin can attack 50-60 of the 358 Lys and Arg residues present in each fibrin molecule, although only 10% are cleaved rapidly. The initial cleavages release the C- and N-termini of the a and b chains, respectively, followed by the cleavages in the extended coiled-coil regions that lead to dissolution of the organized fibrin structure (see Figure 1).
1. Fibrin Polymerization
The generation of fibrin from fibrinogen and its polymerization essentially occur in three sequential steps.
1. Thrombin cleaves the N-terminal regions of the Aa and Bb chains of fibrinogen at the peptide bonds Arg16–Gly17 and Arg14–Gly15, respectively, releasing fibrinopeptides A and B. No further covalent modification to the fibrin(ogen) structure is necessary for polymerization to proceed efficiently. The new N-terminal sequence of the fibrin a chain (Gly17–Pro–Arg–Val) is the major determinant of fibrin polymerization; synthetic peptides corresponding to this sequence can inhibit the polymerization of fibrin monomers (4). By contrast, the new b-chain N-terminal sequence (Gly15–His–Arg–Pro), which is generated more slowly, cannot inhibit polymerization, even though it binds to fibrinogen and to fragment D.
2. The new N-termini of the fibrin monomers spontaneously interact with binding sites in the two homologous distal globular domains, with the g-chain site particularly implicated (3). Both sites are large cavities that bind the a-chain-like peptide Gly–Pro-Arg, but the g-chain cavity is the more specific and does not bind g-chain Gly–His–Arg peptides. It is also the more distal of the two C-terminal domains and is able to associate intermolecularly in a specific end-to-end manner (2). Thus adjacent g-chain sites are closely positioned for optimal binding of the dimeric a-chain N-terminal residues. The efficiency of polymerization derives from both a–g lateral and g–g linear interactions. The symmetry of fibrin(ogen) allows oligomerization to proceed in both directions, generating a two-molecule thick protofibril with a half-staggered overlap (Fig. 1).
3. The fibrin fibrils are reinforced by the transglutaminase activity of factor XIIIa. The g–g linear interactions are the major sites for the introduction of intermolecular isopeptide crosslinks between g-glutamyl carboxyl and lysine-e-amino groups. Subsequently, the globular C-terminal domains of the a chains of adjacent laterally aligned protofibrils are also crosslinked, resulting in the mature, covalently stabilized fibrin mesh. Under physiological conditions, these fibers are of the order of 1 µm in diameter, and thus several hundred fibrin units wide.
2. Dissolution of the Fibrin Clot
The assembly of fibrin also makes accessible epitopes that are involved in its subsequent dissolution by plasmin, after its activation from plasminogen by tissue-type plasminogen activator (tPA). The activity of tPA is potentiated specifically by fibrin, and conformational changes in the vicinity of the C-terminal globular domains of fibrin are thought to reveal the binding sites that are responsible. These sites are residues 312–324 in the C-terminal globular domain of the g chain and 148–160 of Aa, a region that interacts with the Bb-chain C-terminal globular domain. The initial binding sites for plasminogen also reside in fragment D, but subsequent plasmin degradation of fibrin uncovers C-terminal lysine residues that act as supplementary sites for plasminogen binding, which thereby propagate plasminogen activation. Once generated, plasmin can attack 50–60 of the 358 Lys and Arg residues present in each fibrin molecule, although only 5 or 6 are cleaved rapidly. The initial cleavages release the C and N termini of the a and b chains, respectively, followed by the cleavages in the extended coiled-coil regions that lead to dissolution of the organized fibrin structure (see Fig. 1).
References
1. W.R. Fowler and H.P. Erickson (1979) J. Mol. Biol. 134, 241–249.
2. G. Spraggon, S.J. Everse and R.F. Doolittle (1997) Nature, 389, 455–462.
3. K.P. Pratt, H.C. Côté, D.W. Chung, R.E. Stenkamp and E.W. Davie (1997) Proc. Natl. Acad. Sci. USA, 94, 7176–7181.
4. Z. Yang, J.M. Kollman, L. Pandi and R.F. Doolittle (2001) Biochemsitry (In Press(5. A.P. Laudano and R.F. Doolittle (1980) Biochemistry, 19, 1013–1019.



|
|
|
|
طبيبة تبدد 5 خرافات رئيسية عن تغذية الأطفال
|
|
|
|
|
|
|
أول شريحة مزروعة في دماغ إنسان تواجه مشكلة.. ماذا حدث؟
|
|
|
|
|
|
وصل إلى الشلل الرباعي أطباء مستشفى الكفيل يعيدون حركة الأطراف لرجلٍ بعد تكسّر عدد من فقراته
|
|
|
|
بمناسبة ذكرى ولادة السيدة فاطمة المعصومة (عليها السلام) شعبة التوجيه الديني النسويّ تقيم مسابقةً تفاعلية للنساء
|
|
|
|
جامعة الكفيل: مشروع المبنى التعليميّ خطوة مهمّة في توسيع إمكانيات الجامعة ومواكبة التطوّرات العلمية
|
|
|
|
شركة الكفيل للاستِثمارات تباشر بحصاد (320) دونماً من محصول الحنطة
|















