آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Codon Usage and Bias |


|
|
Read More
Date: 22-3-2021
Date: 14-12-2015
Date: 16-5-2016
|
Codon Usage and Bias
The genetic code is degenerate, in the sense that (except for methionine and tryptophan) a given amino acid can be specified by either two, three, four, or six alternative codons. When the same amino acid is specified by three, four, or six codons, they cannot all be recognized by the same transfer RNA anticodon, and so in these cases at least two or three distinct transfer RNAs must carry the same amino acid (isoaccepting) but with different anticodons.
Not all codons for a particular amino acid are used equally frequently. Generally, strong biases favor some codons and are against others, and different codons are favored in different organisms (Table 1). Some genes within an organism show much greater codon usage bias than others, and there is a strong correlation between the degree of bias shown by a gene and its level of expression in terms of the amount of encoded protein produced. For example, in all organisms where the matter has been examined, the genes that encode ribosomal proteins, which are always relatively abundant, are among the most highly biased. The excellent correlation in Escherichia coli between codon bias and level of protein product is shown in Fig. 1. This suggests that the preferred codons are more efficiently translated than the relatively rarely used codons.
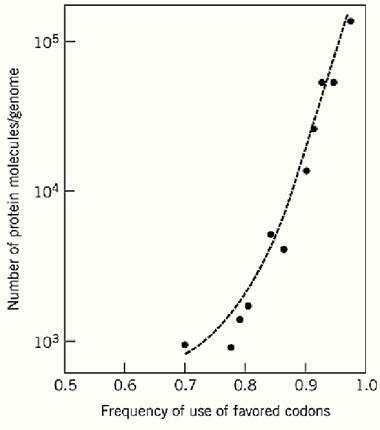
Figure 1. Frequency of using favored (“optimal”) codons in E. coli genes plotted against the quantity of the gene protein products.
Table 1. Codon Bias in Different Organisms a
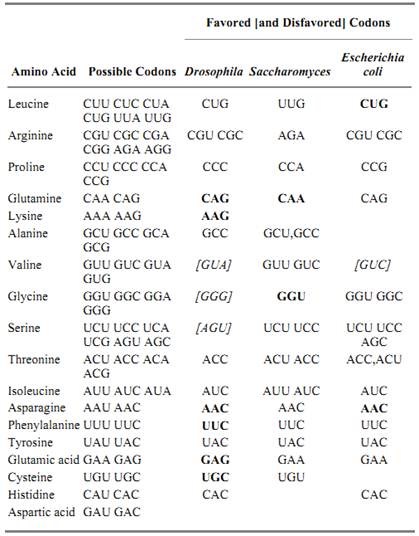
a Drosophila data from a pool of 15 high-expression genes (3); E. coli and Saccharomyces from pools of highly expressed genes (1, 2). The codons indicated show strong majority usage (heavy type indicates 90%) except those italicized in brackets where the bias is strongly against. Blanks indicate no strong bias or insufficient data.
Different translation efficiencies of different codons is largely accounted for by a number of factors, pointed out by Ikemura (“Ikemura's rules”), the first of which is that when different isoaccepting tRNAs are present at very different concentrations, the favored codons will be those that are serviced by the most abundant tRNAs. There is evidence for this generalization from E. coli, Saccharomyces (1, 2) and Drosophila (3). Where a particular amino acid has both codon bias and unequal concentrations of isoaccepting tRNAs, the rule holds virtually without exception. For example, the favored arginine codons in E. coli are CGU and CGC, whereas AGA is strongly favored in Saccharomyces. In the former organism, the tRNA that has anticodon GCI (I for inosine( recognizing CUC/U is present at several-fold higher concentration than that recognizing AGA, with anticodon UCU* (U* is a modified uridine), whereas in yeast the tRNA inequality is very much the opposite.
The different relative abundances of tRNAs do not, however, explain codon usage bias when it operates between codons served by the same isoaccepting tRNA. Here the bias can be explained in terms of Ikemura's other rules, which account for the fact that an anticodon may pair more comfortably with one codon than with another, even though it recognizes both. Thus an anticodon that has a thiolated uridine or 5-carboxymethyl uridine in the 5′ position (the “wobble” pairing position, see Anticodon) prefers a 3′ A in the codon over 3′ G (rule 2); 5′ inosine (I) prefers U or C to A, even though it pairs with all three (rule 3); and codon-anticodon interactions dependent on the relatively weak A–U or T–A pairings in both of the first two positions work best with a stronger G–C or C–G pairing in the third (rule 4). Examples from E. coli consistent with these rules are the biases in favor of GAA over GAG (rule 2), CGU or CGC over CGA (rule 3) and AUC over AUU (rule 4) (1).
1. Effects of Altering Codons
If the more rarely used codons really are translated relatively slowly, one would expect that their introduction into normally highly expressed genes would decrease protein yields. This prediction has been tested experimentally, and substantial effects have indeed been found, especially when multiple and contiguous rare codons have been introduced into the upstream ends of open reading frames. Hoekema et al. (4) carried out a total replacement of the normal codons by synonymous infrequently used codons in the upstream third of the normally highly expressed Saccharomyces PGK1 ) phosphoglycerate kinase) gene and found a tenfold reduction in enzyme yield. There was also a threefold reduction in the level of messenger mRNA. In the gene encoding glutamate dehydrogenase, in Neurospora crassa, a double frameshift mutant that generated three successive rare codons, numbers 54 to 56 of a 453-codon sequence, brought about a threefold reduction in enzyme without appreciable effect on mRNA level (5). And in E. coli, the insertion of an additional mRNA segment containing five contiguous rare AGG arginine codons 24 bases downstream of the initiation codon of lacZ (b-galactosidase) reduced the enzyme yield by 90% in the early exponential phase and virtually to zero thereafter. Another insertion, identical except that AGG was replaced by the favoured CGU codon (6), had little effect. However, when the distance between the inserted AGG-containing sequence and the initiation codon was increased by additional insertions, the yield of b-galactosidase increased almost linearly with distance, as if the rare codons have maximum effect when translation is just starting and progressively less effect as it proceeds. The mechanism of such distance-dependence is unclear. Chen and Inouye (6), reviewing the positions of rare codons in E. coli genes, noted that they fall within the first 25 codons after the initiating AUG and suggested that their function is to modulate the rate of translation. This would only be a regulating mechanism if there were some means of increasing or decreasing the effect, for instance, by adjusting the levels of the least abundant tRNAs. But there is as yet no evidence for this.
If rare codons have strong negative effects on the protein yields of genes, they may be an obstacle to the expression of transgenes in alien host cells. A codon that is abundant in the donor organism may be rare in the recipient. Therefore, to obtain a good yield of a foreign protein, it may be necessary to adjust the coding sequence of the introduced gene by in vitro DNA manipulation. Perhaps the best example of successful application of this idea is provided by the work of Cormack et al. (7), who wished to confer on the fungus Candida albicans the ability to produce the green fluorescent protein) GFP) native to the jellyfish Aequoria victoria. Introduction of the natural GFP gene into Candida on a replicating plasmid vector resulted in the formation of some GFP mRNA in the fungus, but no detectable GFP. Replacing the single CUG codon with CUU, in case the Candida coding idiosyncracy of translating CUG as serine rather than leucine (8) was responsible for the failure, brought no improvement. But when all the Aequoria codons that are comparatively rare in Candida were replaced by codons that are common in the fungus, introduction of the transgene produced an abundance of GF protein. The level of GFP mRNA was also considerably increased, which suggests, as does the yeast PGK1 example mentioned before (4), that blocked translation, and hence failure to cover the mRNA with ribosomes, may expose the mRNA to enzymic degradation.
References
1.T. Ikemura (1985) Mol. Biol. Evol. 2, 13–34.
2. P. M. Sharp, T. M. F. Tuohy, and K. R. Mosurski (1986) Nucleic Acids Res. 14, 5125–5143.
3. D. C. Shields, P. M. Sharp, D. G. Higgins, and F. Wright (1988) Mol. Biol. Evol. 5, 704–716.
4. A. Hoekema, R. A. Kastelein, M. Vasser, and H. A. de Boer (1987) Mol. Cell. Biol. 7, 2914–2924.
5.J. A. Kinnaird and J. R. S. Fincham (1991) J. Mol. Biol. 221, 733–736.
6. G.-F. T. Chen and M. Inouye (1990) Nucleic Acids Res. 18, 1465–1473.
7. B. P. Cormack et al. (1997) Microbiology 143, 303–311.
8. M. A. S. Santos and M. F. Tuite (1995) Nucleic Acids Res. 23, 1481–1486.



|
|
|
|
4 أسباب تجعلك تضيف الزنجبيل إلى طعامك.. تعرف عليها
|
|
|
|
|
|
|
أكبر محطة للطاقة الكهرومائية في بريطانيا تستعد للانطلاق
|
|
|
|
|
|
|
العتبة العباسية المقدسة تبحث مع العتبة الحسينية المقدسة التنسيق المشترك لإقامة حفل تخرج طلبة الجامعات
|
|
|