آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Multiple Regions in RNA Polymerase Directly Contact Promoter DNA |


|
|
Read More
Date: 17-5-2016
Date: 12-5-2016
Date: 17-11-2020
|
Multiple Regions in RNA Polymerase Directly Contact Promoter DNA
KEY CONCEPTS
- The structure of σ70 changes when it associates with core enzyme, allowing its DNA-binding regions to interact with the promoter.
- Multiple regions in σ70 interact with the promoter.
- The α subunit also contributes to promoter recognition.
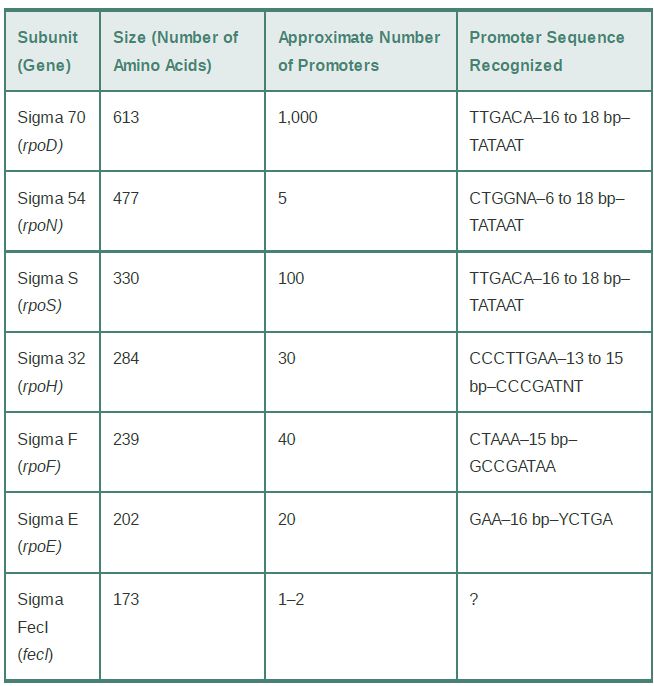
As mentioned briefly in the section titled Sigma Factor ControlsBinding to DNA by Recognizing Specific Sequences in Promoters, several domains in the sigma factor subunit and the CTD in the α subunit of the RNA polymerase core contact promoter DNA. The identification of a series of different consensus sequences recognized by holoenzymes containing different sigma factors (as shown in TABLE 1) implies that the sigma factor subunit must itself contact DNA. This suggests further that the different sigma factors must bind similarly to core enzyme so that the DNA recognition surfaces on the different sigma factors would be positioned similarly to make critical contacts with the promoter sequences in the vicinity of the −35 and −10 sequences.
TABLE 17.1 E. coli sigma factors recognize promoters with different consensus sequences.
Further evidence that sigma factor contacts the promoter directly at both the −35 and −10 consensus sequences was provided by substitutions in the sigma factor that suppressed mutations in the consensus sequences. When a mutation at a particular position in the promoter prevents recognition by RNA polymerase, and a compensating mutation in sigma factor allows the polymerase to use the mutant promoter, the most likely explanation is that the relevant base pair in DNA is contacted by the amino acid that has been substituted.
Comparisons of the sequences of several bacterial sigma factors suggested conserved regions in E. coli σ70 (FIGURE 1) that interact directly with promoters, and these inferences were substantiated by the identification of a crystal structure of RNA polymerase holoenzyme in complex with a promoter fragment. The bacteria T. aquaticus and T. thermophilus illustrate how the DNAbinding regions of the sigma factor fold into independent domains in the protein regions 1.2, 2.3–2.4, 3.0, and 4.1–4.2.
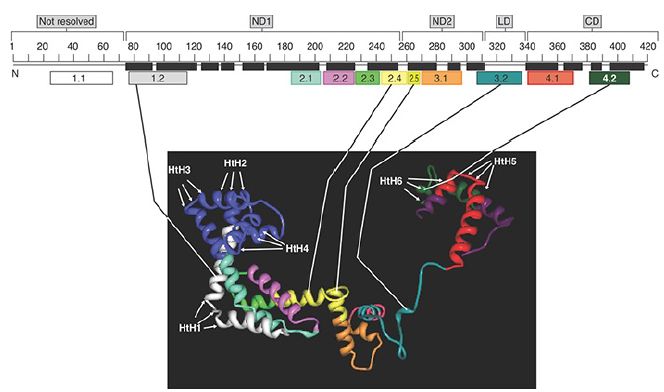
FIGURE 1 The structure of sigma factor in the context of the holoenzyme: −10 and −35 interactions. Sigma factor is extended and its domains are connected by flexible linkers.
Illustration adapted from D. G. Vassylyev, et al., Nature 417 (2002): 712–719. Structure from Protein Data Bank 1IW7.
Figure 1 illustrates the sections of sigma factor that play direct roles in promoter recognition. This figure shows the structure of the major sigma factor as it exists in the context of the holoenzyme. Two short parts of region 2 and one part of region 4 (2.3, 2.4, and 4.2) contact bases in the −10 and −35 elements, respectively; sigma factor region 1.2 contacts the promoter region just downstream from the −10 element, and region 3.0 contacts the promoter region just upstream from the −10 element. Each of these regions forms short stretches of α-helix in the protein. A crystal structure of the holoenzyme in complex with a promoter fragment, in conjunction with experiments with promoters in which the DNA strands were built to contain mismatches (heteroduplexes), showed that σ70 makes contacts with bases principally on the nontemplate strand of the −10 element, the extended −10 element, and the discriminator region, and it continues to hold these contacts after the DNA has been unwound in this region. This confirms that sigma factor is important in the melting reaction.
The use of α-helical motifs in proteins to recognize duplex DNA sequences is common . Amino acids separated by three to four positions lie on the same face of an α-helix and are therefore in a position to contact adjacent base pairs. FIGURE 2 shows that amino acids lying along one face of the 2.4 region α-helix contact the bases at positions −12 to −10 of the −10 promoter sequence.
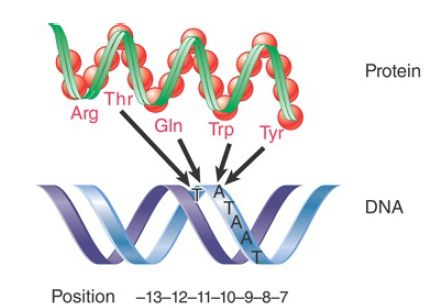
FIGURE 2 Amino acids in the 2.4 α-helix of β70 contact specific bases in the coding strand of the −10 promoter sequence.
Region 2.3 resembles proteins that bind single-stranded nucleic acids and is involved in the melting reaction. Regions 2.1 and 2.2 (which comprise the most highly conserved part of sigma factor) are involved in the interaction with the core enzyme. It is assumed that all sigma factors bind the same regions of the core polymerase, which ensures that the sigma factors compete for limiting core RNA polymerase.
Although sigma factor has domains that recognize specific bases in promoter DNA, the N-terminal region of free sigma factor (region 1.1), acting as an autoinhibitory domain, masks the DNA-binding region; only once the conformation of the sigma factor has been altered by its association with the core enzyme can it bind specifically to promoter sequences (FIGURE 3). The inability of free sigma factor to recognize promoter sequences is important: If sigma factor could bind to promoters as a free subunit, it might block holoenzyme from initiating transcription. Figure 17.17 schematizes the conformational change in sigma factor at open complex formation.
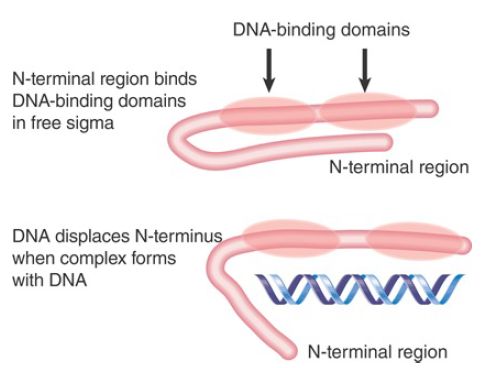
FIGURE 3. The N-terminus of sigma blocks the DNA-binding regions from binding to DNA. When an open complex forms, the Nterminus swings 20 Å away, and the two DNA-binding regions separate by 15 Å.
When sigma factor binds to the core polymerase, the N-terminal domain swings approximately 20 Å away from the DNA-binding domains, and the DNA-binding domains separate from one another by about 15 Å, presumably to acquire a more elongated conformation appropriate for contacting DNA. Mutations in either the −10 or −35 sequences prevent an N-terminal–deleted σ70 from binding to DNA, which suggests that σ70 contacts both sequences simultaneously. This fits with the information from the crystal structure of the holoenzyme (Figure 1), in which it is clear that the sigma factor has a rather elongated structure, extending over the approximately 68 Å of two turns of DNA.
Although sigma factor region 1.1 is not resolved in the crystal structure, biophysical measurements of its position in the holoenzyme versus the open complex suggest that in the free holoenzyme the N-terminal domain (region 1.1) is located in the main DNA channel of the enzyme, essentially mimicking the location that the promoter will occupy when a transcription complex is formed (FIGURE 4). When the holoenzyme forms an open complex on DNA, the N-terminal sigma factor domain is displacedfrom the main channel. Its position with respect to the rest of the protein is therefore very flexible; it changes when sigma factor binds to core enzyme and again when the holoenzyme binds to DNA. The DNA helix has to move some 16 Å from its initial position in order to enter the main DNA channel, and then it has to move again to allow DNA to enter the channel during open complex formation. FIGURE 5 illustrates this movement, looking incross section down t he helical axis of the DNA.
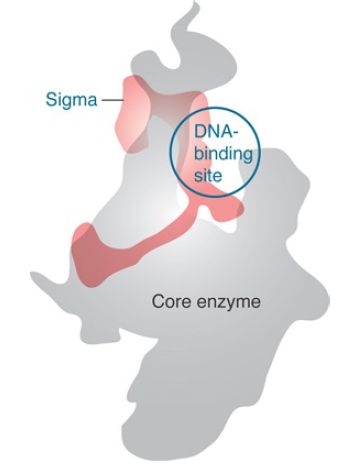
FIGURE 4 Sigma factor has an elongated structure that extends along the surface of the core subunits when the holoenzyme is formed.
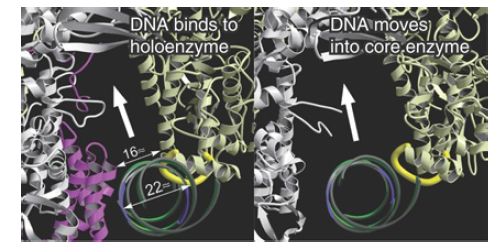
FIGURE 5 DNA initially contacts sigma factor (pink) and core enzyme (gray). It moves deeper into the core enzyme to make contacts at the −10 sequence. When sigma is released, the width of the passage containing DNA increases.
Reprinted by permission from Macmillan Publishers Ltd: Nature, D. G. Vassylyev, et al., vol.417, pp. 712–719, copyright 2002. Photo courtesy of Shigeyuki Yokoyama, The University of Tokyo.
Although it was first thought that sigma factor is the only subunit of RNA polymerase that contributes to the promoter region, the CTD of the two α subunits also can play a major role in contacting promoter DNA by binding to the near promoter UP elements. Because the αCTDs are tethered flexibly to the rest of RNA polymerase , the enzyme can reach regions quite far upstream while still bound to the −10 and −35 elements. The αCTDs thereby provide mobile domains for contacting transcription factors bound at different distances upstream from the transcription start site in different promoters.



|
|
|
|
إجراء أول اختبار لدواء "ثوري" يتصدى لعدة أنواع من السرطان
|
|
|
|
|
|
|
دراسة تكشف "سببا غريبا" يعيق نمو الطيور
|
|
|
|
|
|
جمعية العميد تعقد اجتماعها الدوريّ لمناقشة أنشطتها العلمية المقبلة
|
|
|
|
قسم الشؤون الفكرية يقيم ندوةً عن أهمّية التداول المعرفي لوفدٍ من محافظة بابل
|
|
|
|
السيد الصافي يؤكد أن العتبات المقدسة تمتلك رؤية في دعم الكفاءات العراقية بمختلف تخصصاتها
|
|
|
|
استعدادًا لدوراته القرآنية الصيفية.. المجمع العلمي يقيم دورة تطويرية لمنتسبيه
|