آخر المواضيع المضافة

 الفاكهة والاشجار المثمرة
نخيل التمر
الحمضيات
الاشجار ذات النواة الحجرية
المحاصيل
المحاصيل البقولية
محاصيل الالياف
محاصيل زيتية
محاصيل الحبوب
محاصيل الخضر
القرعيات
المحاصيل المنبهة و المحاصيل المخدرة
المحاصيل السكرية
نباتات الزينة والنباتات الطبية والعطرية
الحشرات النافعة
النحل
دودة القز(الحرير)
تقنيات زراعية
التصنيع الزراعي
الانتاج الحيواني
الطيور الداجنة
الاسماك
الابقار والجاموس
الاغنام
آفات وامراض النبات وطرق مكافحتها
الحشرات
امراض النبات ومسبباتها
الحشائش والنباتات الضارة
الفاكهة والاشجار المثمرة
نخيل التمر
الحمضيات
الاشجار ذات النواة الحجرية
المحاصيل
المحاصيل البقولية
محاصيل الالياف
محاصيل زيتية
محاصيل الحبوب
محاصيل الخضر
القرعيات
المحاصيل المنبهة و المحاصيل المخدرة
المحاصيل السكرية
نباتات الزينة والنباتات الطبية والعطرية
الحشرات النافعة
النحل
دودة القز(الحرير)
تقنيات زراعية
التصنيع الزراعي
الانتاج الحيواني
الطيور الداجنة
الاسماك
الابقار والجاموس
الاغنام
آفات وامراض النبات وطرق مكافحتها
الحشرات
امراض النبات ومسبباتها
الحشائش والنباتات الضارة | التنظيم الجينومي الفايروسي |


|
|
أقرأ أيضاً
التاريخ: 21-6-2018
التاريخ: 13-6-2018
التاريخ: 1-8-2017
التاريخ: 29-6-2018
|
يقصد بالتنظيم الجينومي Genome Organization معرفة التعاقب النيوكليوتايدي Genome Sequence للجينوم الفايروسي وتركيبه وكيفية ترتيب المناطق التشفيرية (الجينات) Coding regions والمناطق غير التشفيرية Non-Coding regions فيه، وأن أول جينوم فايروسي عرف تعاقبه النيوكليوتايدي هو جينوم فايروس موزائيك القرنابيط (CaMV) في سنة 1980، ثم عرف التعاقب النيوكليوتايدي لجينوم فايروس موزائيك التبغ (TMV) سنة 1982، وبحلول سنة 1990 عرف التعاقب النيوكليوتايدي لما يقرب من 40 نوعا من الفايروسات ثم ازدادت إلى 250 نوعا فايروسيا سنة 2000 وشملت أنواعا ممثلة لكل الأجناس الفايروسية تقريبا، كما عرف أيضا التعاقب النيوكليوتايدي الجزئي لجينومات عدد من أنواع الفايروسات وخاصة تعاقب جينات الكابسيد وأفاد ذلك كثيرا في تصنيف الفايروسات وخاصة تمييز أجناس عائلتي Caulimoviridae و Luteoviridae، وتعد دراسة التنظيم الجينومي مفيدة جدا لدراسة حالات التطفير الفايروسية Mutagenesis وتشخيص نواتج الجينات Gene products والمناطق الجينومية غير التشفيرية.
1. الخواص العامة لجينومات فايروسات النبات
يضم الجينوم الفايروسي الجينات وهي المناطق التشفيرية المتمثلة بالتعاقبات النيوكليوتايدية المسئولة عن الإظهار الوراثي Expression للبروتينات اللازمة لإكمال دورة الإصابة الفايروسية وحركة الفايروس في النبات وارتباطه بالناقل، وتضم المناطق التشفيرية أيضا "تعاقبات السيطرة" Control sequences وهي التعاقبات التي تنظم عمل الجينوم، كما يضم الجينوم المناطق غير التشفيرية المسيطرة على تضاعف الفايروس وإظهار الجينوم. يمكن للتعاقبات النيوكليوتايدية الجينومية نظريا تشفير لغاية 12 متعدد الببتيد Polypeptide أو أكثر بقليل، وتشفر معظم الفايروسات الرايبية مفردة الخيط موجبة التوجه (+) ssRNA بين 4-7 أنواع من البروتينات.
توجد الهياكل التشفيرية المفتوحة Open Read Frames , ORFs في المناطق التشفيرية ويعرف "الهيكل التشفيري المفتوح" بأنه تعاقب نيوكليوتايدي يبدأ من "كودون الابتداء" Initiation Codon وهو الكودون AUG (أي تعاقب القواعد: الأدينين واليوراسيل والكوانين وتنتهي بكودون الإنهاء Termination Codon و تشفر عديد ببتيد واحد، وإذا أمتلك الهيكل التشفيري المفتوح إشارة إنهاء مرنة Leaky termination signal عندها يمكن لهذا الهيكل إنشاء عديد ببتيد ثاني. يتراوح عدد الجينات في فايروسات النبات بين جين واحد كما في الفايروس التابع لفايروس موت التبغ (STNV) الى 12 جين كما في بعض انواع الجنسين Closterovirus و Reovirus.
2. كفاءة استعمال الفايروسات لأحماضها النووية
تمتاز الفايروسات بقدرتها العالية على الاستعمال الكفء للكميات المحدودة التي تمتلكها من الأحماض النووية ولكي يحقق الفايروس النباتي هذا الاقتصاد في استعمال حامضه النووي فقد حصلت التحويرات الجينومية التالية في الجينومات الفايروسية (1) دمج التعاقبات التشفيرية بشكل وثيق مع المناطق غير التشفيرية الموجودة في الجينات بأعداد قليلة (2) تراكب التعاقبات التشفيرية لجينين مختلفين في هيكل تشفيري مستقل كما في فايروس الاصفرار التماوتي لعروق البنجر (BNYVV)، (الشكل 1) (3) احتواء جين واحد بالكامل في جين آخر موجود في هيكل تشفيري مختلف كما في فايروس التقزم الأصفر للشوفان ( (OCSV، (الشكل 2) استعمال الية "القراءة الضمنية المتداخلة" Read-through لكودون انهاء طرفي لين Leaky termination Codon لتكوين أكثر من عديد ببتيد واحد وهذه الآلية شائعة مع فايروسات ssRNA(+)، ويمكن تحقيق النتيجة ذاتها بالية أخرى هي "بروتينات التحول الهيكلي" Frame shift proteins والتي يتجنب فيها الرايبوسوم التعامل مع كودون الإيقاف Stop Codon بتحوله إلى هيكل تشفيري آخر مما يسبب تخليق أكثر من عديد ببتيد واحد (4) قيام منتج جيني معين بأكثر من وظيفة فمثلا يقوم الكابسيد لفايروسي موزائيك الجت (AMV) وتخطط الذرة (MSV) بثلاث وظائف الأولى هي حماية الجينوم والثانية نقل الفايروس بالناقل أو نقل الفايروس بين الخلايا كما في الفايروس الثاني والثالثة هي بدء الإصابة من خلال إظهار الأعراض والسيطرة على تضاعف الجينوم (5) وجود الانترونات الوظيفية Functional introns في العديد من فايروسات Geminiviruses وفايروسي تدهور الرز الباسيلي (RTBV) وموزائيك القرنابيط ويعني ذلك حدوث ظاهرة جينومية تسمى "الإقران التراكبي للرنا المراسل" mRNA splicing أي ربط قطعتين جينوميتين بقطعة إضافية وهي العملية التي تزيد من تنويع نسخ الرنا المراسل mRNA transcripts عليه ستزداد أعداد وأنواع منتجات الجينات وهي صفة شائعة في الفايروسات ذات جينوم الدنا (6) قد يستفيد إنزيم التضاعف الفايروسي من أحد بروتينات العائل ليترافق مع ببتيد يشفره الفايروس، وهذا ما لوحظ عند تضاعف فايروس موزائيك التبغ TMV)) (7) تراكب أو تداخل الوظائف التنظيمية الجينومية مع الوظائف التشفيرية أي تراكب التعاقبات التشفيرية مع غير التشفيرية ومثال ذلك إشارات تخليق الجينومات الرايبية الثانوية Subgenomic RNAs لفايروس موزائيك التبغ (8) اشتراك تعاقب نيوكليوتايدي معين في التعاقبات غير التشفيرية في الطرفين 3 و 5 من الجينوم في الفايروسات الرايبية مفردة الخيط لغرض إنجاز أكثر من وظيفة، فقد تبرز هذه التعاقبات في الطرف 5 وهو موقع تعرف الجينوم على الرايبوسومات Ribosome recognition Site وهو أيضا موقع تعرف لأنزيم الربلكيز على النهاية 3 للخيط السالب لبعض الفايروسات. ولوحظ أن الأحماض النووية الفايروسية عموما ينقصها وجود "الأنترونات" Introns حيث وجدت في قلة من الفايروسات النباتية.
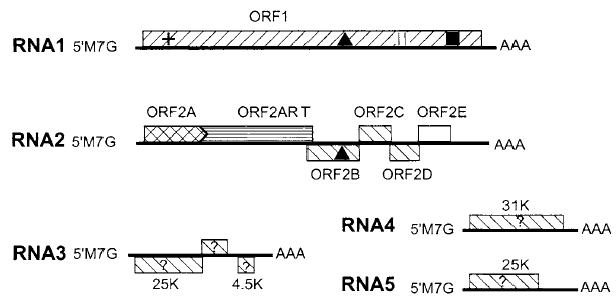
الشكل (1): التنظيم الجينومي لفايروس الاصفرار التماوتي لعروق البنجر (BNYVV) الممثل لفايروسات Benyviruses حيث تمثل الخطوط الكاملة الغامقة قطاعات الجينوم الرايبي المفرد الموجب بقطعه الخمسة المنفصلة، وتدل الصناديق على الهياكل التشفيرية المفتوحة (ORFs) ومنتجاتها الجينية.
مفتاح منتجات الهياكل التشفيرية المفتوحة
الشكل مقتبس منHull ((2002.
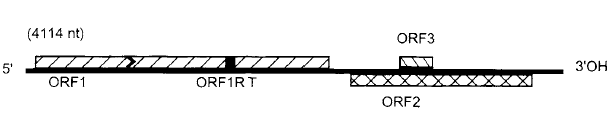
الشكل (2): التنظيم الجينومي لفايروس التقزم الأصفر للشوفان (OCSV) ذو الجينوم الرايبي المفرد الموجب وحيد القطعة، وتدل الصناديق على الهياكل التشفيرية المفتوحة.
مفتاح منتجات الهياكل التشفيرية المفتوحة
.png)
الشكل مقتبس من Hull (2002).
3. وظائف منتجات الجينات الفايروسية
تقوم المنتجات الجينية (البروتينات) التي تشفرها الجينات الفايروسية بالوظائف التالية:
1) الوظائف التركيبية
هي الوظائف التي تنجزها البروتينات التركيبية وتشمل بروتينات الكابسيد في الفايروسات البسيطة وبروتينات الحشوة Matrix واللب Core والبروتينات المرتبطة بالحامض النووي والأغشية الليبوبروتينية في الفايروسات المغلفة وفي عدد من أنواع فايروسات Reoviruses و Tenuiviruses.
2) الوظائف الحيوية
تنجز البروتينات الفايروسية التي تشفرها الجينومات الفايروسية الوظائف الحيوية التالية:
أ- تضاعف الجينوم الفايروسي
هي الوظيفة التي تنجزها إنزيمات تضاعف واستنساخ الجينومات الفايروسية حيث تشفر كل الفايروسات النباتية باستثناء بعض الفايروسات التابعة واحد أو أكثر من أنواع الإنزيمات المبلمرة للحامض النووي Polymerases وهذه الإنزيمات هي مادة حفازة Catalyze قادرة على استنساخ حامض رايبي جديد من حامض رايبي أبوي، ومثالها إنزيم التضاعف الشائع في الفايروسات ذات الجينوم ssRNA(+) وهو "إنزيم بلمرة الرنا المعتمد على الرنا" RdRp والقادر على استنساخ قطع جينومية رايبية مراسلة ثانوية Subgenomic RNAs من الجينوم الأبوي ويطلق على هذه الإنزيم عندما يرتبط مع أغلفة عدد من الفايروسات المغلفة التابعة لفايروسات Rhabdoviruses و Reoviruses بإنزيم "الاستنساخ" Transcriptase، أما إنزيم الربلكيز Replicase فهو الإنزيم القادر على تخليق نسخ جينومية كاملة رايبية من الجينوم الأبوي فضلا عن قدرته على تخليق قطع جينومية رايبية مراسلة ثانوية أقل حجما من الجينوم الأبوي الكامل وقد يتحول هذا الإنزيم إلى معقد إنزيمي تمتد وظائفه إلى ما هو أبعد من نشاط البلمرة ليظهر نشاطا لإنزيمي "الهيليكيز" Helicase " و "المثيل ترانسفيريز" Methyl transferase، وقد سجل هذا مع عدد من الفايروسات حيث لوحظ وجود عدة مناطق وظيفية في هذا المعقد الإنزيمي. يسمى الإنزيم الذي تشفره فايروسات عائلة Caulimoviridae والقادر على استنساخ جينوم كامل الطول من الجينوم الأبوي "الإنزيم المستنسخ العكسي" Reverse transcriptase والذي يسمى أيضا "إنزيم بلمرة الدنا المعتمد على الرنا" RNA dependent DNA polymerase وهو الإنزيم القادر على استنساخ جينوم الدنا من الرنا. وجد في عدد من فايروسات النبات أن للبروتين التركيبي دورا في تضاعف الجينوم الفايروسي حيث يعمل البروتين الفايروسي VP9 عمل البادى Primerr أو يسمى Promote للبدء بتضاعف الجينوم بدءا من النهاية 5 للجينوم ووجد أيضا أن كابسيد فايروسات موزائيك الجت (AMV) وعدد من أنواع فايروسات Ilarviruses يلعب دورا أساسيا في بدء تضاعف الخيط السالب القالب من الخيط الأبوي الموجب.
ب- حركة الفايروسات عبر الخلايا والنقل بالناقلات
تشفر جينومات العديد من الفايروسات بروتينات الحركة لنقل الجسيمات الفايروسية من خلية لأخرى مجاورة عبر البلازموديزماتا كما تساهم هذه البروتينات في الحركة الجهازية للفايروسات في الأوعية الناقلة للنباتات، كذلك تشفر جينومات الفايروسات المنقولة بالناقلات بروتينات متخصصة لربط جسيمة الفايروس بأغشية الناقل وهي المسئولة عن تخصصية النقل.
ج- التعرف على خلية العائل
تمتلك القلة من فايروسات النبات بروتينات متخصصة للتعرف على خلايا العائل Host Cell - recognizing proteins وهي المسئولة عن ربط الجسيمات الفايروسية بالأغشية الخلوية المستهدفة أي بمواقع استقبال على هذه الأغشية تمهيدا لبدء الإصابة الفايروسية حيث تتوفر أدلة متزايدة الآن على مشاركة الأغشية الخلوية في تضاعف الفايروسات، إن معظم فايروسات النبات لا تحتاج التعرف على خلايا النبات لتصيبها لذا فهي ليست بحاجة إلى هذه البروتينات ولكن قد تحتاج إليها الفايروسات المنقولة بالحشرات بالطريقة الباقية لعبور الحواجز داخل أنسجة الناقل والمتمثلة بجدر الأمعاء والغدد اللعابية وفق "الية الاستعانة بالمستقبل" Receptor - mediated mechanism.
4. المناطق غير التشفيرية في الجينوم الفايروسي
تضم المناطق غير التشفيرية Non-coding regions عدة تعاقبات لا علاقة لها بعملية تشفير البروتينات الفايروسية أي أنها ليست جزءا من الجينات التي يحملها الفايروس وفيما يأتي وصفا لهذه المناطق:
1) التراكيب الخاتمة
التراكيب الخاتمة End Structures هي التعاقبات النيوكليوتايدية عند النهايتين 3 و 5 لجينوم والتي تسيطر على عمليتي الترجمة والتضاعف حيث تنظم عمل الهياكل التشفيرية المفتوحة ORFs عند النهاية 5 والمسئولة عن بدء عملية الترجمة، أما التعاقبات التي عند النهاية 3 فتمثل موقع بدء تخليق الخيط السالب القالب.
2) المناطق بين الجينية
المناطق بين الجينية Intergenic regions هي التعاقبات النيوكليوتايدية في المناطق بين الجينات والتي تساهم بتخليق الحامض الرايبي RNA وأيضا في ترجمة الهياكل التشفيرية سفلية الاتجاه Downsream ORFs حيث يبدأ استنساخ القطع الجينومية الرايبية المراسلة الثانوية Subgenomic RNAs من الجينوم الأبوي والتي تعمل بمثابة أنواع من الهياكل التشفيرية المفتوحة بعيدا عن النهاية 5 في العديد من أنواع الفايروسات.
5. التنظيم الجينومي للفايروسات ذات الجينوم الرايبي منقوص الأكسجين مزدوج الخيط dsDNA viruses
درس التنظيم الجينومي لهذه الفايروسات التي تنتمي لعائلة Caulimoviridae والتي تمتاز بجينوماتها الحلقية مع وجود فجوة Disccontinuity or Gap في أحد الخيطين الأصليين الموجبين ووجود فجوتين في الخيط السالب المكمل. يتكون الجينوم من 6-8 جينات في حالة متراكبة مع الجين الثامن، ويبين الشكل (3) ترتيب الهياكل التشفيرية المفتوحة (ORFs) في جينومات عدد من فايروسات هذه العائلة.
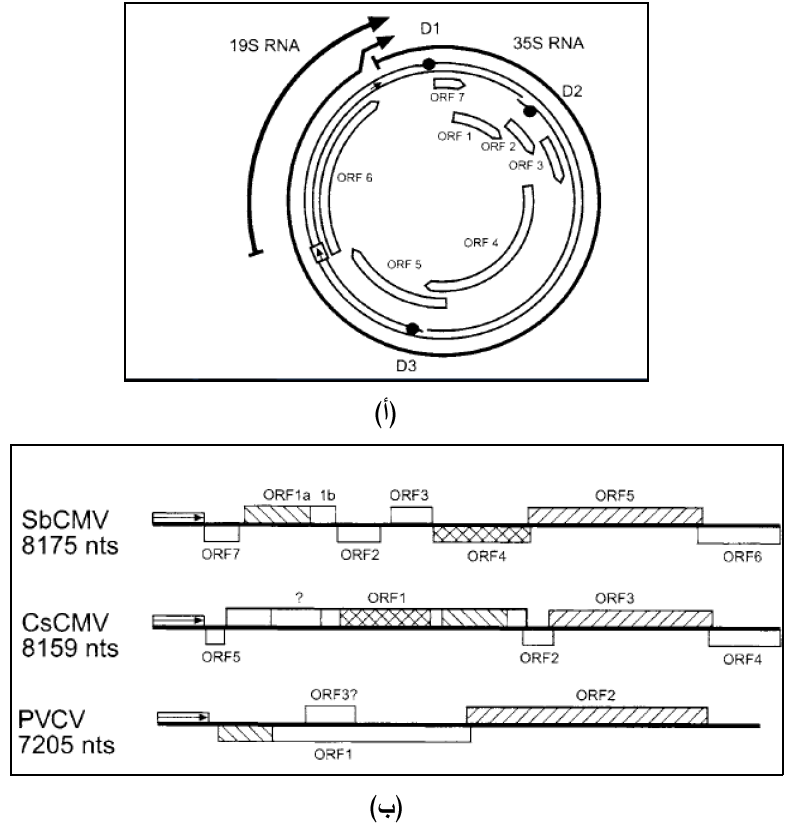
الشكل (3): التنظيم الجينومي لفايروسات من عائلة. Caulimoviridae
(أ) الجينوم الحلقي الفايروس موزائيك القرنابيط (CaMV) حيث تمثل الدوائر السوداء مواقع الفجوات D1 و D2 و D3 فيما تبين الصناديق الحاوية على الأسهم مواقع البواديء Promoters، وتمثل الصناديق الهلالية الطويلة داخل الدائرة الهياكل التشفيرية المفتوحة ORFs السبعة ويشير السهمين الغامقين خارج الدائرة إلى مواقع استنساخ نسخ الحامض الرايبي RNA transcripts وهما 35S و 19S.
(ب) الجينومات الخيطية لثلاثة فايروسات هي التبرقش الأصفر لفول الصويا (SbCMV) والموزائيك العادي للكاسافا (CSCMV) ووضوح عروق البتونيا (PVCV) حيث يبدأ الجينوم بالمنطقة بين الجينية Intergenic region المؤشرة بالأسهم، وتوضح الصناديق مواقع الهياكل التشفيرية المفتوحة أما السهم العمودي فيوضح موقع الفجوة D1، والارقام يمين الشكل تمثل حجم كل جينوم مقاسا بوحدة nt - نيوكليوتايدة.
المفتاح لمنتجات الهياكل التشفيرية
الشكل مقتبس منHull (2002).
6. التنظيم الجينومي للفايروسات ذات الجينوم الرايبي منقوص الأكسجين مفرد الخيطssDNA viruses
درس التنظيم الجينومي للجنسين التابعين لعائلة Geminiviridae الذي يمتازان بالجينوم الحلقي ذو القطعة واحدة أو القطعتين والذي يمتاز بالتعاقب الجينومي المحافظ Conserved genome Sequence، ويوجد في الجينوم منطقة بين جينية غير تشفيرية كبيرة تسمى "المنطقة المشتركة" Common region والتي تمتاز بالتعاقب النيوكليوتايدي المحافظ وهو TAATATTAC والموجود في كل فايروسات هذه العائلة. أعتمد تصنيف أنواع هذه الفايروسات على صفات تنظيمها الجينومي ، وفيما يأتي وصفا له في أجناس هذه العائلة:
1) الجنس Mastrevirus
درس التنظيم الجينومي لفايروس تخطط الذرة (MSV) ذو الجينوم الحلقي المفرد بطول 2687 نيوكليوتايدة وهو جينوم لا يشابه كثيرا جينومات بقية أفراد هذه العائلة التوأمية حيث توجد فيه منطقتين بين جينيتين وهما "المنطقة الطويلة", LIR Large intergenic regionوالمنطقة القصيرة Short intergenic region, SIR وأربع مناطق تشفيرية محتملة، (الشكل 4) وربما يبدأ الاستنساخ من المنطقة الطويلة وينتهي في القصيرة، ويوجد هيكلين تشفيريين ORFs على الخيط الفايروسي الأصلي Viral Strand , V حيث يشفر الهيكل التشفيري ORFV1 بروتين الحركة عبر الخلايا ويشفر الهيكل التشفيري ORFV2 الغطاء البروتيني، ويوجد هيكلين آخرين على الخيط المكمل (C) Complementary Strand وهما ORF C1 و ORF C2 واللذان يشفران البروتينات المشاركة في التضاعف.
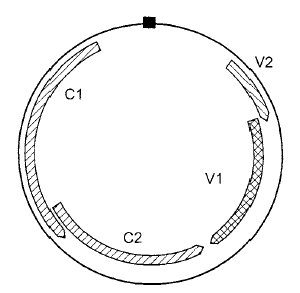
الشكل (4): التنظيم الجينومي لجينوم فايروس تخطط الذرة (MSV) الحلقي نوع ssDNA. يمثل الصندوق الأسود أعلى الدائرة المنطقة المشتركة أما الصناديق الهلالية داخل الدائرة فتمثل الهياكل التشفيرية المفتوحة (ORFs).
المفتاح لمنتجات الهياكل التشفيرية

الشكل مقتبس منHull 2002)).
2) الجنس Begomovirus
أنواع هذا الجنس ذات جينوم ثنائي القطعة Bipartite genome وتتباين التعاقبات النيوكليوتايدية في كلا القطعتين باستثناء المنطقة المشتركة غير التشفيرية والتي بطول 200 نيوكليوتايدة، وجد أن معظم فيروسات البيجومو المنتشرة في القارات القديمة والتي تسمى "فايروسات بيجومو العالم القديم" Old World begomoviruses هي ذات جينوم ثنائي القطعة أما فايروسات بيجومو العالم الجديد"New world begomoviruses المنتشرة في الأمريكيتين فكلها ثنائية القطعة حيث توجد 5-6 جينات على القطعة الأولى وجين واحد أو أثنين على الخيط الموجه بالفريون Virion Sense Strand وأربعة جينات على الخيط المكمل Complementary sense strand , AC القطعة الثانية فتحمل جينين يوجد الأول وهو BV1 على الخيط الموجه بالفريون فيما يوجد الأخر وهو BC1 على الخيط المكمل، يشفر الهيكل ORF AV1 وحدات الغطاء البروتينية ويشفر الهيكل ORF AV2 بروتين الحركة عبر الخلايا فيما تشارك البروتينات التي تشفرها الجينات المحمولة على الخيط المكمل للقطعة A في عمليتي التضاعف والإظهار، ويشفر الهيكل ORF AC1 إنزيم التضاعف، فيما يشفر الهيكل ORF AC2 بروتين منشط لترجمة البروتينات، ويشفر الهيكل ORF AC3 بروتينات معززا للتضاعف Replication enhancer protein ولا تعرف وظيفة الهيكل ORF AC4، فيما يشفر الهيكل ORF BV1 الموجود في القطعة الجينومية الثانية (B) "بروتين الوشيعة النووية" Nuclear Shuttle protein، ويشفر الهيكـل ORF BC1 بروتينا يساعد في حركة الفايروس عبر الخلايا، أما التنظيم الجينومي لفايروسات "البيجومو" أحادية القطعة فهو مماثل تماما لتنظيم القطعة الأولى في الفايروسات ثنائية القطعة، (الشكل 5).
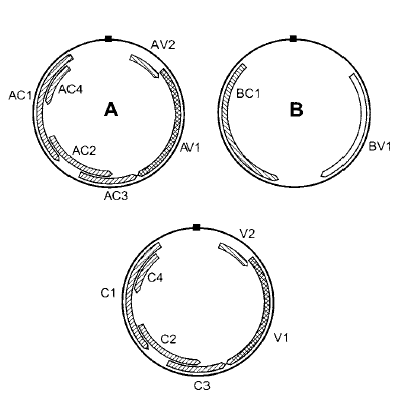
الشكل (5): التنظيم الجينومي لفايروسات Begomovirus. تمثل الدائرتين العلويتين الفايروسات ثنائية القطعة الجينومية، فيما تمثل الدائرة السفلى جينوم الفايروسات أحادية القطعة الجينومية. يمثل الصندوق الأسود في أعلى كل دائرة، المنطقة المشتركة، أما الصناديق الهلالية داخل الدوائر فتمثل الهياكل التشفيرية المفتوحة (ORFs).
المفتاح لمنتجات الهياكل التشفيرية
الشكل مقتبس من Hull (2002).
7. التنظيم الجينومي للفايروسات ذات الجينوم الرايبـي مزدوج الخيط dsRNA viruses
درس التنظيم الجينومي للجنسين التابعين لعائلة Reoviridae واللذان يضمان أنواعا من الفايروسات ذات الجينوم الرايبي المزدوج متعدد القطع Multiple linear dsRNA والتي تتراوح بين 10-12 قطعة وترقم وفقا لحركتها الكهربية في الهلام Electrophoretic mobility وأبطأها حركة هي القطعة الأولى 1 Segment ثم تتوالى بقية القطع وهي جميعها أحادية التشفير Monocistronic أي أن كل قطعة تشفر بروتينا واحدا لأنها تحوي هيكلا تشفيريا (ORF) واحدا ولكن وجدت بعض القطع ثنائية التشفير لامتلاكها هيكلين تشفيرييين، تشفر هذه الفايروسات عدة منتجات جينية هي بروتينات الغطاء وإنزيم التضاعف وبروتينات أخرى لا تعرف وظيفتها ووجد أن هناك تماثل في التنظيم الجينومي لهذه الفايروسات مع مثيله في فايروسات تصيب الفقريات وتعود لنفس العائلة وذلك في بعض مواقع الجينوم.
1) الجنس Fijivirus
يقسم جينوم أنواع هذا الجنس إلى عشرة قطع ويوجد تماثل جينومي مع جينوم الفايروس الحشري المنتمي لنفس الجنس وهو الفايروس NLRV) Nilaparavata lugens reovirus) ويفسر التماثل الجينومي بين الفايروسات النباتية والحشرية في هذا الجنس إلى تضاعف الأولى في أجسام ناقلاتها الحشرية.
2) الجنس Phytoreovirus
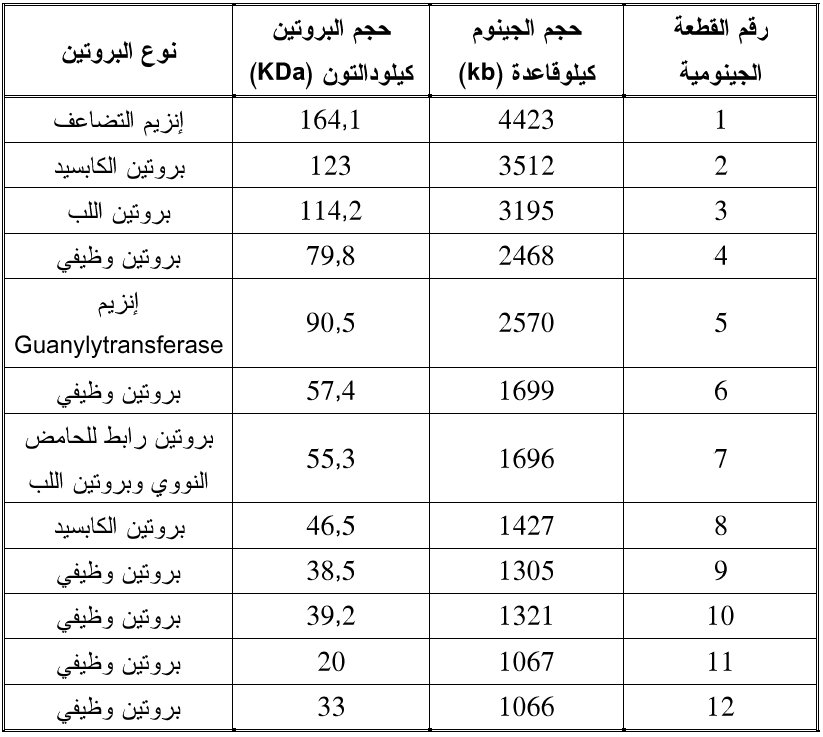
يقسم جينوم أنواع هذا الجنس إلى 12 قطعة وأكثر الفايروسات دراسة هو فايروس تقزم الرز (RDV) والذي يلخص الجدول (1) خواص جينومه وحجوم ووظائف البروتينات الفايروسية التي يشفرها.
الجدول (1): خواص جينوم فايروس تقزم الرز (RDV) ووظائف قطعه الاثنا عشر
8. التنظيم الجينومي للفايروسات ذات الجينوم الرايبي مفرد الخيط سالب التوجه ss RNA viruses (-)
تنتمي هذه الفايروسات لعائلتي Rhabdoviridae و Bunyaviridae اللتان تضمان فيروسات مغلفة وكذلك إلى الجنسين المستقلين Tenuivirus و Ophiovirus حيث يشفر جينوم هذه الفايروسات العديد من أنواع البروتينات الفايروسية أهمها إنزيم التضاعف RNA polymerase المرتبط بالفريون والمحمول فيه والذي يعد ضروريا للاستنساخ الابتدائي للجينوم الأبوي.
1) عائلة Rhabdoviridae
تمتلك أنواعها جينوما من قطعة واحدة بطول 11-13 ألف نيوكليوتايدة، ودرس التعاقب النيوكليوتايدي لفايروس الاصفرار التماوتي للخس (LNYV) والفايروس الشبكي الأصفر للجعضيض (SYNV) المتشابهين كثيرا في تنظيمهما الجينومي، (الشكل 6) حيث يوجد في النهاية 3 تعاقب نيوكليوتايدي يسمى "التعاقب الريادي leader Sequence مكون من 144 نيوكليوتايدة مع مناطق بين جينية قصيرة بينها، ويشفر الجين N بروتينا بحجم 54 كيلودالتون مرتبط تركيبيا مع الجينوم، اما الجين G فيشفر البروزات السكرية البروتينيه Glycosylated Spikes بحجم 70 كيلودالتون تمتد عبر الغلاف الفايروسي إلى سطح الجسيمة، ويشفر الجين L إنزيم بلمرة الرنا" بحجم 241 كيلودالتون المنطمر في الفريون، ويشفر الجين SC4 البروتين 37 كيلودالتون المرتبط بالغلاف الفايروسي والغني بالحامضين الأمينيين "السيرين" Serine و"الثريونين" Threonine والذي يعتقد أنه يسيطر على تضاعف الفايروس أو يسهل حركته عبر الخلايا.
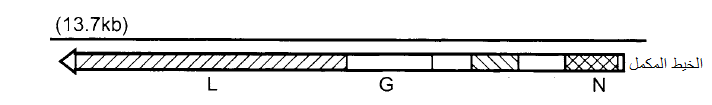
الشكل (6): التنظيم الجينومي لفايروس SYNV. يمثل الخط الأسود المتصل العلوي، الجينوم الأبوي السالب، أما السهم الأسفل فيشير إلى الخيط الموجب المكمل والذي يحمل المناطق التشفيرية (الجينات) ومنتجاتها البروتينية حيث يشير رأس السهم إلى النهاية 5.
المفتاح لمنتجات الهياكل التشفيرية
الشكل مقتبس من Hull (2002).
9. التنظيم الجينومي للفايروسات ذات الجينوم الرايبي مفرد الخيط موجب ss RNA viruses Alaan (+)
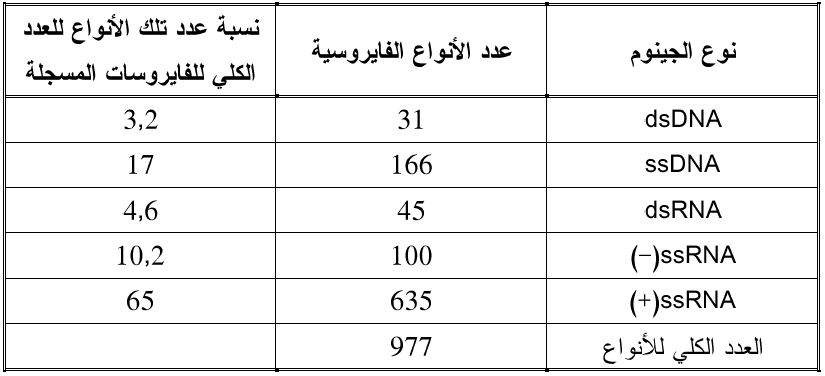
تمتلك حوالي 65% من مجمل الأنواع الفايروسية المسجلة فايروسات النبات جينوما من النوع SS RNA(+) بشكل قطعة واحدة أو مقسم إلى عدة قطع، (الجدول 2).
الجدول (2): أنواع الجينومات الفايروسية حسب توزيعها في الأنواع الفايروسية المسجلة
وفيما يلي وصف للتنظيم الجينومي لأكبر عائلتين في هذه المجموعة من الفايروسات
1) عائلة Potyviridae
يتكون جينوم خمسة من الأجناس التابعة لهذه العائلة من قطعة جينومية واحدة Monopartite genome باستثناء الجنس Bymovirus فهو ذو جينوم ثنائي القطعة Bipartite genome ويمتاز جينوم هذه العائلة بأنه عديد الأدينيل Polyadenylated ويرتبط معه البروتين 5VPg من النهاية 5 للجينوم وهو بوزن جزيئي 24 كيلودالتون مع وجود منطقة غير تشفيرية في النهاية 5 بحجم 144 نيوكليوتايدة غنية بالقاعدتين النتروجينيتين "الأدنين واليوراسيل" ووجود هيكل تشفيري كبير (ORF) بطول 9161 نيوكليوتايدة يشفر عديد ببتيد بطول 300 حامض أميني وبوزن جزيئي 340 كيلودالتون، ويمتاز أيضا بوجود منطقة جينومية غير قابلة للترجمة عند النهاية3 untraslated region 3بطول 190 نيوكليوتايدة تنتهي بقاعدة الأدنين المأذنلة Poly A، يشفر هذا الجينوم بروتين كبير واحد Polyprotein والذي ينشطر إلى 9 أو 10 قطع بروتينية بواسطة إنزيم تفكيك البروتين Proteinase الذي يشفره الجينوم الفايروسي أيضا وتقوم كل منها بوظيفة معينة، يبين الشكل (8) التنظيم الجينومي لفايروس نقش التبغ (TEV) والبروتينات التي يشفرها الجينوم والتي تؤدي وظائف حيوية عديدة للفايروس ومنها القطعة التي بوزن جزيئي 30 كيلودالتون والبروتين VP9 اللذان يدخلان في بناء الكابسيد وبروتين "المكون المساعد" Helper Component protein المعروف اختصارا HC-Pro وهو المسئول عن النقل التخصصي للفايروس بحشرات المن حيث يشكل عامل الربط بين كابسيد الفايروس وأنسجة المن لذلك فهو بروتين نقل" ويحتوي على عنصر الزنك والبروتين NIa الذي يمثل إنزيم البروتينيز والبروتين Nib الذي يمثل إنزيم البوليميريز RdRP.
2) عائلة Bromoviridae
تمثل هذه العائلة الفايروسات النباتية الرايبية مفردة الخيط متعددة القطع الجينومية، وأكثرها دراسة هو فايروس موزائيك نجيل البروم (BMV) الذي يمتاز بالجينوم ثلاثي القطع Tripartite genome بطول 8243 نيوكليوتايدة، (الشكل 8) حيث تشفر القطعة RNA1 عديد ببتيد بوزن جزيئي 110 كيلودالتون فيما تشفر القطعة RNA2 عديد ببتيد بوزن جزيئي 105 كيلودالتون وتشفر القطعة RNA3 عديدي ببتيد بوزن جزيئي 20 و 35 كيلودالتون على التوالي وتشفر القطعة RNA4 بروتين الغطاء وهو بوزن جزيئي 20 كيلودالتون ولوحظ أن ترجمة قطعة الرنا الرابعة RNA4 تكون أكفأ بوجود القطع الجينومية الثلاثة الأولى وربما يفسر ذلك بزيادة كفاءة الارتباط بالرايبوسوم وتساهم القطعتين RNA1 و RNA2 في تشفير إنزيم الربلكيز ولهاتين القطعتين قدرة التضاعف لوحدهما دون القطع الأخرى كما يلعب البروتين المشفر من قبل القطعة الثالثة دورا في حركة الفايروس عبر الخلايا.
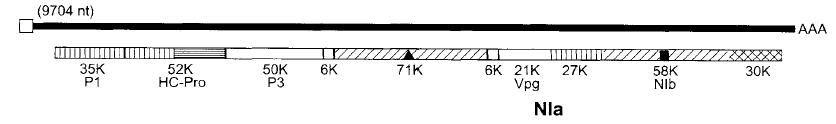
الشكل (7): التنظيم الجينومي لفايروس نقش التبغ (TEV). الخط الأسود العلوي يمثل الجينوم الفايروسي المفرد وعدد نيوكليوتايداته، فيما يوضح الخط الأسفل المناطق التشفيرية (الصناديق المظللة) وأنواع البروتينات التي تشفرها.
المفتاح لمنتجات الهياكل التشفيرية
المصدر: (2002) Hull.
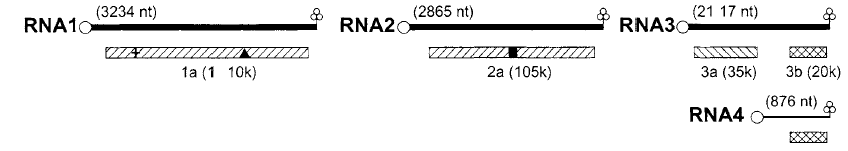
الشكل (8): التنظيم الجينومي لفايروس موزائيك نجيل البروم (BMV) حيث تمثل الخطوط الغامقة العلوية الثلاثة القطع الجينومية وعدد نيوكليوتايداتها، اما الخط الرفيع السفي فيمثل القطعة الجينومية الرابعة، وتلاحظ الصناديق اسفل الخطوط والتي تمثل منتجات الجينات.
المفتاح لمنتجات الهياكل التشفيرية

الشكل مقتبس من Hull (2002).
ويبين الشكل (9) التنظيم الجينومي لفايروس موزائيك الجت (AMV) والمكون جينومه من اربع قطع هي RNA1 و RNA2 و RNA3 و RNA4.
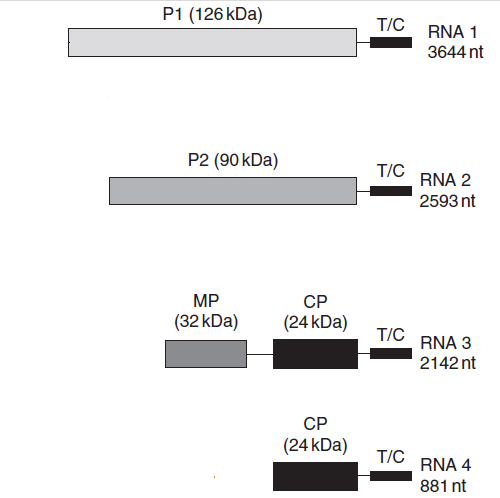
الشكل (9) التنظيم الجينومي لفايروس موزائيك الجت (AMV).
تشفر القطعتين RNA1 و RNA2 بروتيني إنزيم الربلكيز P1 وP2 وتشفر القطعة RNA3 بروتين الحركة MP وبروتي الكابسيد CP الذي تشفر جزءا منه أيضا القطعة RNA4، تمثل القطعة T/C جزءا من الجينوم بحجم 112 نيوكليوتايدة (nt) والتي تماثل في تركيبها الرنا المراسل Trna حجوم البروتينات التي تشفرها هذه القطع مبينة بالكيلو دالتون.
الشكل مقتبس من Mahy و Van Regenmortel (2008).



|
|
|
|
"عادة ليلية" قد تكون المفتاح للوقاية من الخرف
|
|
|
|
|
|
|
ممتص الصدمات: طريقة عمله وأهميته وأبرز علامات تلفه
|
|
|
|
|
|
|
قسم التربية والتعليم يكرّم الطلبة الأوائل في المراحل المنتهية
|
|
|