آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Deoxyuridine Triphosphatase |


|
|
Read More
Date: 11-12-2020
Date: 14-3-2021
Date: 9-12-2020
|
Deoxyuridine Triphosphatase
Living cells contain two enzyme systems whose primary role is evidently to assure the exclusion of uracil from DNA where thymine occurs instead. Deoxyuridine triphosphatase (dUTPase) catalyzes the hydrolysis of dUTP to deoxyuridine monophosphate and pyrophosphate, thereby removing from the nucleotide pool a precursor, dUTP, that is recognized by DNA polymerases as readily as is thymidine triphosphate for incorporation into DNA opposite template adenine residues. The other system involves a DNA base excision repair process that begins with the action of DNA-uracil N-glycosylase. This enzyme cleaves the glycosidic bond linking uracil to deoxyribose, and an oligonucleotide patch that contains the abasic site is removed and replaced.
Current interest in dUTPase focuses on its very high substrate specificity, its significance as a possible chemotherapeutic target, the evolution and functions of virus-coded forms of dUTPase, and the generation and regulation of organelle-specific isoforms in eukaryotic cells.
1. Biological Functions of dUTPase
Although dUTPase was discovered in the early 1960s (1, 2), the enzyme was not characterized as a highly purified protein until 1978 (3). Much of the interest in the 1970s related to the role of uracil metabolism in creating short DNA fragments that had been identified as intermediates in DNA replication. After Okazaki proposed that DNA replication involves the discontinuous synthesis of short DNA fragments—which became known as Okazaki Fragments—several laboratories reported that nascent DNA chains are complementary to both template DNA strands, suggesting that both daughter DNA strands grow discontinuously. However, Tye et al. (4) showed that many of the short fragments arose not as intermediates in replication but through the action of the uracil exclusion systems. Incorporation of dUMP into DNA, followed by the action of uracil N-glycosylase and base excision repair, caused DNA strand breakage that converted nascent high-molecular-weight DNA into small fragments. When corrections were made for this factor, it was found that leading-strand DNA replication occurs continuously, with short fragments of DNA arising as replication intermediates only in synthesis of the lagging strand.
Why uracil should be excluded from DNA was not immediately apparent because uracil is identical to thymine in its base pairing with adenine, and so its substitution for thymine was not expected to affect DNA function or genetic specificity significantly. Indeed, the existence of bacteriophages, such as the Bacillus subtilis phage PBS2, whose DNA contains uracil but no thymine (5), indicates that substituting U for T is biologically acceptable. However, it was recognized that uracil can arise in DNA either through incorporation of dUMP opposite dAMP or through deamination of dCMP in a G · C base pair. This latter event is potentially mutagenic because the resultant G · U base pair would be converted to an A · T base pair in subsequent rounds of replication. Because dUTPase has no direct involvement in these events, it was thought that dUTPase should be a dispensable function in most organisms. However, El-Hajj et al. (6) were unable to isolate a null mutant of dUTPase in Escherichia coli, leading them to speculate that the protein plays an additional, essential metabolic role; similar results were reported for yeast (7). In a later study, however, El-Hajj et al. (8) analyzed a conditional lethal dUTPase (dut) mutation of E. coli and found that it could be suppressed by a mutation in dcd, the structural gene for dCTP deaminase, the enzyme that synthesizes dUTP in enteric bacteria. This finding, plus the construction of a multiple mutant bacterium that survived extensive substitution of uracil for thymine in its DNA (8), indicated that the primary role of dUTPase is indeed to keep the dUTP pool small and facilitate the exclusion of uracil from DNA. The lethality of dUTPase null mutants is ascribed, then, to excessive accumulation of dUMP in DNA, leading to double-strand breaks that result when repair of single-strand breaks occurs at nearby sites on opposite DNA strands.
2. Structure of Deoxyuridine Triphosphatase
The dUTPase molecule must be constructed to exacting design specifications in order to cleave efficiently one deoxyribonucleoside triphosphate while having little or no effect on structurally similar dNTPs needed for DNA synthesis, in particular, dUTP and dTTP. Accordingly, kinetic analysis of E. coli dUTPase (9) showed the ratio kcat/Km to be, remarkably, more than105-fold lower for dCTP than for dUTP. The nucleotides dTTP and UTP were even poorer substrates as no action of dUTPase on these nucleotides was detectable. The effects of pH on the Km for dUTP suggested important contributions of hydrogen bonding involving N3 and the carbonyl oxygen on C4 in establishing the specificity of dUTP binding to dUTPase, which is consistent with structural studies (10, 11). Crystallographic analysis of both the E. coli and human enzymes reveals a homotrimeric structure, with each of three substrate-binding sites made up from residues in two adjacent polypeptide chains. The bacterial enzyme was crystallized in the presence of a competitive inhibitor, dUDP, which identified the active site and provided a structural explanation of the kinetic observations.
3. dUTPase as a Chemotherapeutic Target
Thymidylate metabolism has long been recognized as a target for drugs that would act to inhibit DNA replication in either cancer or infectious diseases. Beginning in the early 1980s, it was recognized that a significant contributor to the lethality of inhibitors of dihydrofolate reductase or thymidylate synthase is the accumulation of dUTP and its resultant incorporation into DNA (12). Inhibition of thymidylate synthase flux either directly, by 5-fluorodeoxyuridylate, or indirectly, by inhibition of dihydrofolate reductase, was seen to cause accumulation of dUMP which, in turn, would be converted to dUTP faster than it could be cleaved by dUTPase. Hence, the resultant accumulation of dUMP in DNA was seen to cause double-strand DNA breaks by the process discussed above. The existence of a class of fluorodeoxyuridine-resistant cell mutants with elevated levels of dUTPase (13) is consistent with this interpretation. Now that X-ray crystallography structures for dUTPase are available, this enzyme has been recognized as a suitable target for structure-assisted drug design. In principle, dUTPase inhibitors could be used, either alone or in conjunction with inhibitors of dihydrofolate reductase or thymidylate synthase, to kill proliferating cells selectively by stimulating the incorporation of dUMP into their DNA.
4. dUTPase Isoforms in Eukaryotic Cells
One of the most intriguing recent developments concerning dUTPase (14) relates to the discovery of two different isoforms in mammalian cells, one located predominantly in the nucleus and the other localized to mitochondria (although an earlier study (15) had localized human dUTPase to the cytosol). Presumably, the mitochondrial isoform plays the same role in mitochondrial DNA synthesis as the nuclear enzyme plays for nuclear DNA replication. Of great interest is the recent finding of Ladner and Caradonna (16) that both isoforms are encoded by the same nuclear gene, with alternative splicing at the 5′ end of the dUTPase gene (Fig. 1). The use of different 5′ exons for the two isoforms gives a subunit size of 22 kDa for the cytosolic enzyme and 23 kDa for the mitochondrial isoform. This may be the first reported instance of alternative splicing of the same transcript to generate isoforms of the same enzyme localized in different cell compartments.
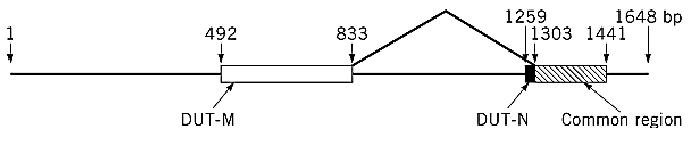
Figure 1. Alternative splicing at the 5′ end of the human dUTPase gene as a means to generate nuclear and mitochondrial isoforms of dUTPase. Exons are boxed, and open reading frames of the mitochondrial, nuclear, and common sequences are shown by different shading. Intron/exon boundaries are indicated and numbered with respect to an arbitrary start point for the 5′ end of the gene. The amino-terminal splice junction is indicated by joined lines.
Cell-cycle analysis of protein and messenger RNA levels shows that the mitochondrial enzyme is expressed constitutively throughout the cell cycle (as is mitochondrial DNA replication), whereas the expression of the nuclear form is closely linked to nuclear DNA replication status. The nuclear isoform undergoes protein degradation as cells exit the cycle. Whether this differential regulation is related to phosphorylation of the enzyme is an intriguing unanswered question. Ladner et al. (14) showed that the cytosolic dUTPase is phosphorylated at Ser11, in a cyclin-dependent protein kinase phosphorylation site, whereas the same site remains unphosphorylated in the mitochondrial isoform. The functional significance of this difference has not yet been defined.
5. Virus-Encoded dUTPases
Numerous virus-specific forms of dUTPase have been described, including those encoded by the RNA genomes of retroviruses. The first viral dUTPase to be described (17) was shown to be an activity of dCTPase, an enzyme induced after T-even bacteriophage infection, which is responsible for exclusion of cytosine from phage DNA. This activity is essential to the observed complete replacement of cytosine by 5-hydroxymethylcytosine in the phage genome. Purification of dCTPase revealed an associated dUTPase activity (18), and both activities were shown to be intrinsic to the same protein. Also, unlike the unrelated bacterial and eukaryotic dUTPases, the phage enzyme is equally active on the respective deoxyribonucleoside diphosphates, cleaving dCDP and dUDP to dCMP and dUMP, respectively, plus inorganic phosphate. In T4, this multifunctional enzyme is encoded by gene 56. Mutants defective in this gene are lethal because they incorporate dCMP into phage DNA, and the viral genome encodes nucleases that specifically recognize and degrade cytosine-containing DNA.
dUTPase is encoded by the genomes of herpes viruses, pox viruses, several nonprimate lentiviruses, and type B and D retroviruses. A sequence in the gag-pro gene region of retrovirus genomes, originally identified as the evolutionary result of the duplication of a proteinase gene, was found, as these sequences began to accumulate in the data banks, to be homologous to dUTPase genes (19). Cloning and expression of these “pseudoproteinase” genes showed that they did indeed express functional dUTPases (20, 21). It is of particular interest that the human immunodeficiency virus ) HIV) genome does not encode a dUTPase, although such a gene is expressed by the closely related feline immunodeficiency virus (FIV), which is being studied as a model for HIV infection. Lerner et al (22) infected cats with wild-type FIV and with a dUTPase-negative FIV mutant. After nine months of infection, sequence analysis of viral genomes isolated from both groups of infected animals showed a fivefold increase in mutations in the dUTPase-negative infections, most of them involving G → A transitions. These could have arisen from the formation of rG · dU base pairs during reverse transcription in the presence of expanded dUTP pools. Of great interest is the possibility that the extreme genetic variability of HIV is related to the lack of a dUTPase gene in the wild-type HIV genome, giving HIV the same mutator phenotype as observed in dUTPase-deficient FIV mutants.
References
1. L. E. Bertani, A. Häggmark, and P. Reichard (1961) J. Biol. Chem. 236, PC67–PC68.
2. G. R. Greenberg and R. L. Somerville (1962) Proc. Natl. Acad. Sci. USA 48, 247–257.
3. J. Shlomai and A. Kornberg (1978) J. Biol. Chem. 253, 3305–3312.
4. B-K. Tye, P-O. Nyman, I. R. Lehman, S. Hochhauser, and B. Weiss (1977) Proc. Natl. Acad. Sci. USA 74, 154–157.
5. A. R. Price and H. R. Warner (1975) J. Biol. Chem. 250, 8804–8811.
6. H. H. El-Hajj, H. Zhang, and B. Weiss (1988) J. Bacteriol. 170, 1069–1075.
7. M. H. Gadsden, E. M. McIntosh, J. C. Game, P. J. Wilson, and R. H. Haynes (1993) EMBO J. 12, 4425-4431.
8. H. H. El-Hajj, L. Wang, and B. Weiss (1992) J. Bacteriol. 174, 4450–4456.
9. G. Larsson, P. O. Nyman, and J-O. Kvassman (1996) J. Biol. Chem. 271, 24010–24016.
10. G. Larsson, L. A. Svensson, and P. O. Nyman (1996) Nature Struct. Biol. 3, 532–538.
11. C. D. Mol, J. M. Harris, E. M. McIntosh, and J. A. Tainer (1996) Structure 4, 1077–1092.
12. M. Goulian, B. Bleile, and B. Y. Tseng (1980) J. Biol. Chem. 255, 10630–10637.
13. C. E. Canman, T. S. Lawrence, D. S. Shewach, H. Y. Tang, and J. Maybaum (1993) Cancer Res. 53, 5219–5224.
14. R. D. Ladner, D. E. McNulty, S. A. Carr, G. D. Roberts, and S. J. Caradonna (1996) J. Biol. Chem. 271, 7745–7751.
15. J. A. Vilpo and H. Autio-Harmainen (1983) Scand. J. Clin. Lab. Invest. 43, 583–590.
16. R. D. Ladner and S. Caradonna (1997) J. Biol. Chem. 272, 19072–19080.
17. G. R. Greenberg (1966) Proc. Natl. Acad. Sci. USA 56, 1226–1232.
18. A. R. Price and H. R. Warner (1969) Virology 39, 882–892.
19. D. J. McGeoch (1990) Nucleic Acid Res. 18, 4105–4110.
20. A-C. Bergman, O. Björnberg, J. Nord, P. O. Nyman, and A. M. Rosengren (1994) Virology 204, 420-424.
21. N. A. Roseman, R. K. Evans, E. L. Mayer, M. A. Rossi, and M. B. Slabaugh (1996) J. Biol. Chem. 271, 23506–23511.
22. D. L. Lerner, P. C. Wagaman, T. R. Phillips, O. Prospero-Garcia, S. J. Henriksen, H. S. Fox, F. E. Bloom, and J. H. Elder (1995) Proc. Natl. Acad. Sci. USA 92, 7480–7484.



|
|
|
|
اكتشاف تأثير صحي مزدوج لتلوث الهواء على البالغين في منتصف العمر
|
|
|
|
|
|
|
زهور برية شائعة لتر ميم الأعصاب التالفة
|
|
|
|
|
|
موكب أهالي كربلاء يستذكر شهادة الإمام الصادق (عليه السلام)
|
|
|
|
العتبة العباسية تستذكر شهادة الإمام الصادق (عليه السلام) بإقامة مجلس عزاء
|
|
|
|
أهالي كربلاء يحيون ذكرى شهادة الإمام الصادق (عليه السلام) في مدينة الكاظمية
|
|
|
|
شعبة مدارس الكفيل النسوية تعقد اجتماعًا تحضيريًّا لوضع الأسئلة الامتحانية
|