آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Cytokeratins |


|
|
Read More
Date: 1-5-2016
Date: 27-12-2015
Date: 11-5-2016
|
Cytokeratins
Cytokeratins form long filaments (they are one of the family of intermediate filament proteins) that extend in a sinuous manner between the cytoplasmic surface of the cell nucleus and the plasma membrane of epithelial cells. The cytokeratin filaments appear to attach to the cytoplasmic side of the nuclear envelope, often close to the nuclear pore complexes. From there they radiate outward and link to or loop through the portions of the desmosomes or adhesion plaques that lie on the inner side of the cell membrane.
Cytokeratin chains can be divided into two groups: “acidic” Type I and “neutral-basic” Type II. In general, the Type II chains have higher isoelectric points and higher molecular weights than their Type I counterparts. Each chain comprises nonhelical end domains separated by a flexible rodlike structure. The N- and C-terminal domains in the epidermal keratins (or cytokeratins) have a characteristic substructure arranged about the rod domain with bilateral symmetry: E1, V1, H1 in the N-terminal domain and H2, V2, E2 in the C-terminal domain (1). The H1 and H2 subdomains, located respectively immediately N- and C-terminal to the rod domain, have sequences that are highly homologous regions within each chain type. The V1 and V2 subdomains are variable in length and sequence but always have a high content of glycine and serine residues. The E1 and E2 subdomains lie at the extreme ends of the molecule and are short, generally basic regions.
Between the terminal domains, the amino acid sequence has a heptad repeat substructure characteristic of a coiled-coil conformation (2). The latter thus consists essentially of coiled-coil rod domains (segments 1A, 1B, 2A, and 2B), joined by short linking domains (L1, L12, and L2), and results in a rod of total length 46.2 nm (Fig. 1). Each molecule contains a pair of different chains) Type I and Type II) aligned parallel to one another and in axial register (3-5). Segments 1B and 2 =) 2A + L2 + 2B) each contain highly regular distributions of both acidic and basic residues (2). Thus each rod segment can be modeled as alternating bands of positive and negative charge, with a period of about 1.4 to 1.5 nm. Pairs of molecules aggregate through their rod domains in three antiparallel modes: (i) with their 1B segments largely overlapped, (ii) with their 2B segments largely overlapped, and (iii) with their entire lengths largely overlapped. In cytokeratins there is also a short head-to-tail overlap of about eight residues between parallel molecules (6). The antiparallel interactions originate in large part by the maximization of electrostatic interactions, which are possible as a consequence of the regular charge distribution in neighboring rod domains. They are also both specified and stabilized by interactions between the H1 (and presumably H2) subdomains, the head-to-tail overlap region, and the link L2. Cross-links formed between lysine residues in cytokeratins have been characterized (Fig. 2), thus confirming the modes of aggregation originally proposed on theoretical grounds. This work also allowed the projected (axial) lengths of the links L1, L12, and L2 to be determined, together with the relative axial staggers between various coiled-coil rod segments (6, 7). These data allow a surface lattice structure to be constructed that predicts a near-axial period in the cytokeratin of 22.6 nm (the experimentally observed value is22.7 nm). Scanning transmission electron microscope (STEM) data indicate that in vitro assemblies may contain about 16, 24, 32, or 40 chains, but it seems that only the 32-chain variant is present in vivo. The STEM data, nonetheless, would indicate that the cytokeratin may have a substructure in which an eight-chain entity, a protofibril, is present. In vitro two, three, four, or five of these protofibrils may aggregate in a nonspecific manner.
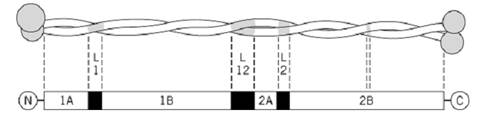
Figure 1. Schematic cytokeratin intermediate filament molecule. The rod domain is of approximate length 46 nm, with coiled-coil segments 1A, 1B, 2A, and 2B and non-coiled-coil links L1, L12, and L2. A break in the phasing of the heptad repeat, known as a stutter, occurs at the center of segment 2B. The N- and C-terminal domains contain E (end), V (variable but rich in glycine and serine residues), and H (homologous) subdomains arranged about the rod domain with bilateral symmetry.
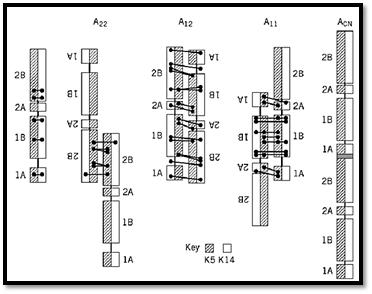
Figure 2. The cross-links induced chemically between lysine residues are shown for human K5/K14 epidermal keratin IF. The cross-links between the two polypeptide chains within a molecule (left) show that the chains are parallel and in axial register. Cross-links between molecules indicate that the molecules aggregate antiparallel through one of three modes, with (i) their 2B segments largely overlapped (A22), (ii) the entire molecules largely overlapped (A12), and/or ) iii) their 1B segments largely overlapped (A11). Combination of A22 and A11 results in a small head-to-tail overlap of about 1 nm between parallel molecules.
Characterization of a large number of gene mutations leading to keratinopathies has been achieved in recent years. The resulting sequence changes lie very largely in or close to the head-to-tail overlap region comprising the conserved sequences at the N-terminal end of segment 1A and the C-terminal end of segment 2B. A reasonable conclusion is that this interaction is subtly modified by the mutated residue and becomes less able to stabilize the cytokeratin in vivo.
References
1. P. M. Steinert, D. A. D. Parry, W. W. Idler, L. D. Johnson, A. C. Steven, and D. R. Roop (1985( Amino acid sequences of mouse and human epidermal type II keratins of Mr 67000 provide a systematic basis for the structural and functional diversity of the end domains of keratin intermediate filament subunits. J. Biol. Chem. 260, 7142–7149.
2. W. G. Crewther, L. M. Dowling, P. M. Steinert, and D. A. D. Parry (1983) Structure of intermediate filaments. Int. J. Biol. Macromol. 5, 267–274.
3. P. M. Steinert (1990) The two-chain coiled-coil molecule of native epidermal keratin intermediate filaments is a type I–type II heterodimer. J. Biol. Chem. 265, 8766–8774.
4. M. Hatzfeld and K. Weber (1990) The coiled-coil of in vitro assembled keratin filaments is a heterodimer of type I and type II keratin: use of site-specific mutagenesis and recombinant protein expression. J. Cell Biol. 110, 1199–1210.
5. P. A. Coulombe and E. Fuchs (1990) Elucidating the early stages of keratin filament assembly. J. Cell Biol. 111, 153–169.
6. P. M. Steinert, L. N. Marekov, R. D. B. Fraser, and D. A. D. Parry (1993) Keratin intermediate filament structure: crosslinking studies yield quantitative information on molecular dimensions and mechanism of assembly. J. Mol. Biol. 230, 436–452.
7. P. M. Steinert, L. N. Marekov, and D. A. D. Parry (1993) Conservation of the structure of keratin intermediate filaments: molecular mechanism by which different keratin molecules integrate into pre-existing keratin intermediate filaments during differentiation. Biochemistry 32K 10046–10056 .
8. D. A. D. Parry and R. D. B. Fraser (1985) Intermediate filament structure: 1, Analysis of IF protein sequence data. Int. J. Biol. Macromol. 7, 203–213.



|
|
|
|
دخلت غرفة فنسيت ماذا تريد من داخلها.. خبير يفسر الحالة
|
|
|
|
|
|
|
ثورة طبية.. ابتكار أصغر جهاز لتنظيم ضربات القلب في العالم
|
|
|
|
|
|
|
العتبة العباسية المقدسة تقدم دعوة إلى كلية مزايا الجامعة للمشاركة في حفل التخرج المركزي الخامس
|
|
|