آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Cytochromes |


|
|
Read More
Date: 4-4-2021
Date: 12-5-2016
Date: 22-3-2021
|
Cytochromes
Cytochromes are iron-containing proteins that facilitate the movement of electrons in a wide variety of metabolic processes. Cytochromes are widely distributed throughout nature. They are found in all eukaryotes and in most, but not all, prokaryotes. The prosthetic group or chromophore of cytochromes is heme, an iron-containing porphyrin. The prototypic form of heme in cytochromes is protoheme IX, which consists of iron, two vinyl side chains, four methyl groups and two propionic acid side chains on a conjugated tetrapyrole ring (Fig. 1) . Protoheme IX is the prosthetic group of b-type cytochromes and the family of proteins known as cytochrome P450. The c-type cytochromes have heme c as the prosthetic group, in which protoheme IX is covalently bound to the protein through one or, more commonly, two thioether bonds to cysteine side-chains. Heme a, found in a-type cytochromes, has a long isoprenoid tail substituted on one of the vinyl groups and a formyl group replacing a methyl. Further variations in heme structure are found in isolated cases, for example, cytochrome d and cytochrome o (1) but will not be discussed here. Cytochrome P450 and cytochrome c are discussed elsewhere in this volume.
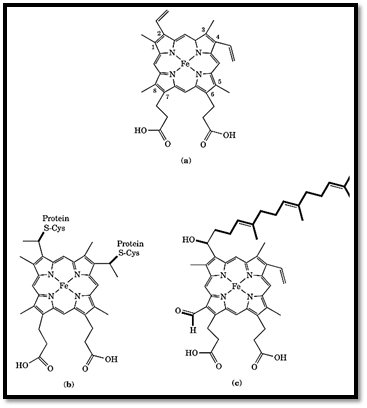
Figure 1. The structures of (a) protoheme IX, (b) heme c, and (c) heme a.
The iron in cytochromes, is either penta- or hexa-coordinate. Porphyrin nitrogen atoms fill four coordination positions (in-plane coordination, see Fig. 1) and appropriate ligands are positioned below the heme plane (5-coordinate) and, in most cases also above the heme plane (6-coordinate). The out-of-plane ligands in typical c-type cytochromes, are the N3 atoms of histidine residues and the sulfur atom of methionine (Fig. 2). Ligation in the b-type cytochromes is generally by two His residues (Fig. 2) and by one or two His in the cytochromes a. Variations exist, however. For example, the out-of-plane ligation is by two His residues in cytochrome c3, and by one His and an open position in cytochrome c′. Cytochromes are associated with the inner membrane of mitochondria, that of microsomes, the thylakoid membrane in chloroplasts, and the plasma membrane in bacteria. However, soluble forms (typically c-type cytochromes) are found in the space between the inner and outer mitochondrial membranes and in the periplasmic space in bacteria.

Figure 2. Stereo representations of the structure of tuna cytochrome c (upper stereo pair) and microsomal cytochrome b5 (lower stereo pair). In cytochrome c, the N-terminal a-helix is at the top, the Met80 ligand is on the left, and the His18 ligand on the right. In cytochrome b5, the N-terminus is at the lower right the His61 ligand is on the left, and the His37 ligand is on the right. The heme propionates are oriented toward the solvent.
The physiological activity in all cytochromes is the reversible oxidation and reduction of the iron atom, which cycles between the ferric (Fe3+) and ferrous (Fe2+) states. From a physiological standpoint, an important property of cytochromes is their oxidation-reduction (redox) potential, which is Eo′ at pH 7. The redox potential measures the relative stabilities of the oxidized (ferric) and reduced (ferrous) forms and therefore their tendencies to donate and accept electrons, respectively. At pH 7 in aqueous solution, the redox potential of H2 is –414 mV and that of O2 is +816 mV. With relatively minor exceptions, these define the biological range. Cytochromes have redox potentials somewhere between these limits. In biological electron transfer, the direction of electron flow is from lower to higher potentials in the limit from hydrogen to oxygen. Thus the redox potential of a particular cytochrome determines, to a first approximation, where it is positioned relative to other redox compounds in electron transfer pathways. The c-type
cytochromes have redox potentials ranging from –400 to +450 mV, depending on the structural family and species, although they typically vary between only +200 and +300 mV in eukaryotic c-type cytochromes. The b-type cytochromes have redox potentials that range from –200 to +150 mV, and the a-type cytochromes have potentials in the +220 to +400 mV range.
The role of the protein moiety in cytochromes is threefold. First, the protein provides the out-of-plane ligands and, in the case of c-type cytochromes, the covalent attachment sites for the heme. Secondly, the protein provides the proximal environment for the heme, a nonpolar molecule that is not water-soluble, controls the exposure of the heme to the solvent, and interacts with the heme to modulate the redox potential. Thirdly, the protein surface provides the sites for the interaction with electron donors and acceptors because in all known cases the heme iron is buried in the protein's interior. Thus, the redox potential is modulated by a combination of the nature of the out-of-plane ligands, the immediate protein environment, and the solvent exposure of the heme.
The redox potentials dictate the direction of electron flow. For example, mitochondrial cytochrome c 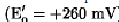 cannot significantly reduce oxidized pyridine nucleotide (NAD+,
cannot significantly reduce oxidized pyridine nucleotide (NAD+,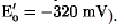 They do not, however, explain the specificity and efficiency of biological electron transfer. Within a biological milieu, a large number of redox compounds exist close to each other. These include small organic compounds, such as flavins and pyridine nucleotides (for example, NAD+); oxygen (in aerobes); inorganics, such as nitrates, nitrites, sulfates, and sulfhydryls; and a variety of redox proteins, including non-heme-iron sulfur proteins, flavoproteins, cytochromes, and copper proteins (see Electron Transfer Proteins). Thus, from an energetic standpoint, substantial opportunities exist for electrons to flow in a great variety of directions. This does not happen, and electron flow occurs with very high efficiency (approaching 100%) only within specific pathways (for example, in respiration or photosynthesis). This incredible efficiency, the essence of life, is mediated by the surface of the protein, which provides recognition sites that determine which redox compounds interact on a physiological timescale, that is, it determines how fast electrons are transferred, and the direction of electron flow (2, 3). This kinetic control means that energetically equivalent reactions can take place on timescales that differ by many orders of magnitude. For example, reduced pyridine nucleotide reduces mitochondrial cytochrome c slowly (in seconds), whereas cytochrome c1
They do not, however, explain the specificity and efficiency of biological electron transfer. Within a biological milieu, a large number of redox compounds exist close to each other. These include small organic compounds, such as flavins and pyridine nucleotides (for example, NAD+); oxygen (in aerobes); inorganics, such as nitrates, nitrites, sulfates, and sulfhydryls; and a variety of redox proteins, including non-heme-iron sulfur proteins, flavoproteins, cytochromes, and copper proteins (see Electron Transfer Proteins). Thus, from an energetic standpoint, substantial opportunities exist for electrons to flow in a great variety of directions. This does not happen, and electron flow occurs with very high efficiency (approaching 100%) only within specific pathways (for example, in respiration or photosynthesis). This incredible efficiency, the essence of life, is mediated by the surface of the protein, which provides recognition sites that determine which redox compounds interact on a physiological timescale, that is, it determines how fast electrons are transferred, and the direction of electron flow (2, 3). This kinetic control means that energetically equivalent reactions can take place on timescales that differ by many orders of magnitude. For example, reduced pyridine nucleotide reduces mitochondrial cytochrome c slowly (in seconds), whereas cytochrome c1 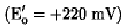
reduces the same cytochrome in milliseconds, even though reduction by pyridine nucleotide is more strongly favored energetically in the first case (65.7 kJ/mol versus 3.8 kJ/mol.( The interactive domains of the cytochromes are defined by the amino acid side chains positioned on their molecular surface that dictate their sites of interaction. The structure of the complex between two electron transfer proteins determines the proximity and orientation of the relevant prosthetic groups (the two hemes in the case of electron transfer between two cytochromes). Three factors play a role in defining the interactive domains. First, the positioning of charged amino acid side chains on the surface of interacting molecules result in electrostatic interactions, either attractive or repulsive, that define the interaction domain (4). Secondly, the distance between redox centers (heme irons in the case of cytochromes) influences the rate of electron transfer. This distance is controlled by the size and nature of the amino acid side chains in the interactive domain (2). Third, two interactive domains that interact must be complementary physically and structurally, even if defined by nonpolar van der Waals interactions (2, 3). Apparently, during the course of evolution, mutations have occurred that optimize the rate of electron flow through relevant metabolic pathways by modulating the amino acid side chains defining the interactive domains, thus allowing competing redox reactions to occur in close proximity and channeling the flow of electrons through the appropriate pathway.
From the standpoint of solubility, cytochromes exist in three types of environments. Soluble forms are typically c-type cytochromes that are free to diffuse between electron donors and acceptors. Integral membrane proteins, that is, transmembrane proteins, have only part of the molecule exposed to solvent. The majority of the molecule is integrated into the membrane. Examples are a-type cytochromes and some b-type cytochromes. Peripheral membrane proteins are strongly associatedwith a membrane or with integral membrane proteins and have a substantial portion of the molecule in solution. Examples are cytochrome b5 and cytochromes c1 and f.
1. A-type Cytochromes
Cytochromes a and a3 are components of cytochrome oxidase, or complex IV, of the mitochondrial electron transfer chain and are found in all eukaryotic organisms and many aerobic prokaryotes. Cytochrome oxidase catalyzes the four-electron reduction of O2 to H2O, utilizing cytochrome c as the electron donor. Cytochrome a, which is hexa-coordinate, mediates the transfer of electrons from cytochrome c to the penta-coordinate cytochrome a3 (with the intermediate participation of a copper atom;), which provides the site for oxygen reduction. Variants of cytochrome oxidase are found in some aerobic bacteria that do not contain hemes other than heme a. Examples are cytochrome bo, which uses quinols as the electron donor in Escherichia coli and cytochrome cbb3 in Rhodobacter sphaeroides, which has cytochrome c as an electron donor (1). Thus, the heme type can vary, although the function, the reduction of molecular oxygen to water, remains constant. Cytochrome oxidase is an integral membrane protein and couples the transfer of electrons and the corresponding redox energy with the movement of protons across the inner mitochondrial or periplasmic membrane, driving the synthesis of ATP. From an evolutionary standpoint, there is structural and amino acid sequence homology between eukaryotic and bacterial cytochrome oxidase (1). In addition, a variety of other enzymes exist that are structurally related to cytochrome oxidase and generally use cytochrome c or an electron donor, such as nitrous oxide reductase, which reduces N2O to N2 and utilizes copper as the prosthetic group, but has sequence homology with cytochrome oxidase and utilizes cytochrome c as its electron donor (1). Another example is nitric oxide reductase, which catalyzes the reduction of nitric oxide to nitrous oxide. Although it is a cytochrome bc complex, it has a transmembrane component and homology to portions of cytochrome oxidase (1).
2. B-type Cytochromes
The b-type cytochromes fall into two broad categories: (1) a family of relatively large cytochromes containing protoheme IX that are part of integral membrane protein complexes like cytochrome bc1 and cytochrome b6f; (2) a family of cytochrome b5-like proteins that are peripheral proteins, typically anchored to a membrane, or are components of protein complexes like flavocytochrome b2 and sulfite oxidase. The cytochrome bc1 complex and its homologue in plants, cytochrome b6f, catalyze the transfer of electrons from quinones to cytochrome c (in animals) or to plastocyanin (in plants) coupled with the movement of protons across the inner mitochondrial or the thylakoid membranes, which drives the synthesis of ATP. The mitochondrial cytochrome bc1 complex, also known as complex III, has the cytochrome b component integrated into the membrane through a number of transmembrane a-helices (5). The cytochrome c1 component is anchored to the membrane and is the electron donor to cytochrome c. Cytochrome b is also a component of succinate dehydrogenase (also known as complex II), a complex integral membrane protein that participates in the mitochondrial electron transfer pathway (6). Succinate dehydrogenase oxidizes succinate to fumarate. The electrons reduce quinone and feed into the mitochondrial electron transfer chain.
Because of their large size and strong association with membranes, cytochromes b are not as well understood as the generally small and water-soluble c-type cytochromes (cytochromes c1 and f are exceptions in terms of size and solubility). Thus the extent of structural and functional diversity among the cytochromes b, particularly in prokaryotes, remains an open question.
Proteins of the cytochrome b5 family contain a cytochrome b5 domain (Fig. 2) and, in the case of the mitochondrial and microsomal members, a hydrophobic tail or anchor that associates the cytochrome with the relevant membrane (7). The structure of cytochrome b5 is shown in Fig. 2. In nitrate reductase, sulfite oxidase, and flavocytochromes b2, the cytochrome b5 domain is tightly associated with a flavoprotein that specifies the particular enzymatic activity. The cytochromes b5 domain acts as an electron acceptor from the molybdenum (Mo)-pterin cofactor in sulfite oxidase or from protein-bound flavin in cytochrome b5 reductase, nitrate reductase, P450 reductase, and flavocytochrome b2. The cytochrome b5 electron acceptor is cytochrome c in the flavocytochrome b2 and sulfite oxidase systems, and a Mo-pterin cofactor in nitrate reductase. In microsomal systems, cytochrome P450 and a variety of acyl-CoA desaturases are the electron acceptors. In red blood cells, cytochrome b5 reduces methemoglobin. Thus, cytochrome b5 has evolved the ability to function in a diverse group of electron transfer pathways and with a variety of electron donors and acceptors (7).
3. C-type Cytochromes
These cytochromes are discussed in more detail elsewhere in this encyclopedia, but it is important to note that this is a diverse group of cytochromes consisting of a variety of structural motifs and functions (8). Although restricted to mitochondrial cytochrome c and cytochrome c1 (discussed previously) in eukaryotes and key components in the respiratory electron transfer chain, a variety of bacterial c-type cytochromes have evolved into a diverse and complex array of structures and function. In terms of structural families, the cytochromes c include the mitochondrial cytochrome c family and their bacterial homologies, cytochrome c1 and f, and the cytochromes c3, cytochromes c′, flavocytochromes c, cytochrome c4, and cytochromes c5. Now, only cytochrome c3 and c-type cytochromes that are related to mitochondrial cytochrome c are well understood in both structure and function.
The basic structural motif of the mitochondrial cytochrome c family has been maintained throughout evolution. Figure 2 presents the structure of tuna cytochrome c. In contrast to cytochrome b5, the heme propionates are buried and not solvent accessible. Cytochromes utilizing the cytochrome c structural motif are found in almost all bacteria but differ primarily in their redox potentials, which vary from approximately +50 to +450 mV, and in the type and distribution of amino acid side chains on their molecular surfaces.
4.Summary
Cytochromes evolved early in the development of life on this planet, and they have retained the basic functional group protoheme IX (with relatively minor variations) throughout evolution. A number of basic structural motifs have evolved that are found in both prokaryotes and eukaryotes, including the mitochondrial cytochromes c and their bacterial precursors, the cytochrome b5 family, the cytochrome oxidases, the membrane-spanning cytochromes b, and the cytochrome c1/b6 family. It is striking that a relatively small number of structural families have evolved to take advantage of diverse metabolic opportunities, capturing redox energy to produce ATP and adapting to available ecological niches.
References
1. M. Saraste, J. Castresana, D. Higgins, M. Lübben, and M. Wilmanns (1996) In Origin and Evolution of Biological Energy Conversion (H. Baltscheffsky, ed.) VCH, New York, pp. 255–289.
2. G. Tollin, T. E. Meyer, and M. A. Cusanovich (1986) Biochim. Biophys. Acta 853, 29–41.
3. T. E. Meyer, G. Tollin, and M. A. Cusanovich (1994) Biochimie 76, 480–488.
4. J. A. Watkins, T. E. Meyer, G. Tollin, and M. A. Cusanovich (1994) Protein Sci. 3, 2104–2114.
5. P. N. Furbacker, G.-S. Tae, and W. A. Cramer (1996) In Origin and Evolution of Biological Energy Conversion (H. Baltscheffsky, ed.), VCH, New York, pp. 221–253.
6. B. A. C. Ackrell, M. K. Johnson, R. P. Gunsalus, and G. Cecchini (1992) In Chemistry and Biochemistry of Flavoenzymes, Vol. III (F. Müller, ed.), CRC Press, Boca Raton, pp. 229–297.
7. F. Lederer (1994) Biochimie 76, 674–692.
8. T. E. Meyer, J. J. van Beeumen, R. P. Ambler, and M. A. Cusanovich (1996) In Origin and Evolution of Biological Energy Conversion (H. Baltscheffsky, ed.), VCH, New York, pp. 71–108.



|
|
|
|
إجراء أول اختبار لدواء "ثوري" يتصدى لعدة أنواع من السرطان
|
|
|
|
|
|
|
دراسة تكشف "سببا غريبا" يعيق نمو الطيور
|
|
|
|
|
|
قسم الشؤون الفكرية يقيم برنامج (صنّاع المحتوى الهادف) لوفدٍ من محافظة ذي قار
|
|
|
|
الهيأة العليا لإحياء التراث تنظّم ورشة عن تحقيق المخطوطات الناقصة
|
|
|
|
قسم شؤون المعارف يقيم ندوة علمية حول دور الجنوب في حركة الجهاد ضد الإنكليز
|
|
|
|
وفد جامعة الكفيل يزور دار المسنين في النجف الأشرف
|