آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Secondary Phloem |


|
|
Read More
Date: 29-10-2015
Date: 9-11-2016
Date: 7-11-2016
|
Secondary Phloem
Because secondary phloem is formed from the vascular cambium just as secondary xylem is, it too has an axial and a radial system (Fig. 1). The axial system is responsible for conduction up and down the stem or root; it contains sieve tube members and companion cells in dicots, or sieve cells in gymnosperms. In both groups of plants, fibers and non-conducting parenchyma are also typically present in axial secondary phloem. In some species, there may be bands of fibers alternating with sieve tube members, but usually these are not annual rings, and many other patterns can be seen in other species. Whereas the equivalent cells of the axial secondary xylem are arranged as early and late wood, and as ring porous or diffuse porous wood, no similar arrangement occurs in secondary phloem. Although the tracheary elements of secondary xylem may function for many years before being converted to heartwood, sieve tube members and sieve cells conduct for less than 1 year; only the innermost layer of phloem is capable of conduction.
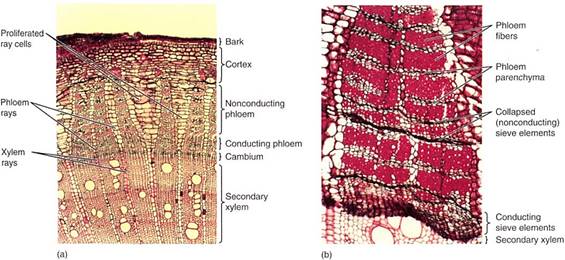
FIGURE 1 :(a) This stem transverse section has secondary xylem at the bottom, vascular cambium region, secondary phloem, then cortex and bark at the top. Notice that the xylem rays and the phloem rays meet at the cambium. Parenchyma cells of some rays have proliferated through cell division and expansion, which has prevented tearing of the bark (X 40). (b) In this secondary phloem of Artabotrys, only the youngest axial phloem (at the bottom) contains functional sieve tube members. The youngest phloem and the vascular cambium are so soft that they are often damaged while trying to cut samples from trees; that is why they are partially crushed here. Older phloem has abundant fibers alternating with bands of phloem parenchyma and collapsed sieve tube members (X 50).
The size, shape, and number of phloem rays match those of xylem rays because both are produced by the same ray initials (Fig. 2). Phloem rays consist only of parenchyma cells that are used for storage, as are xylem rays, but phloem rays seem to be even more important for this (Fig. 3). In gymnosperms, albuminous cells are ray cells.
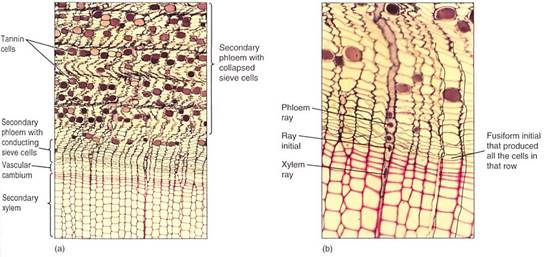
FIGURE 2 (a and b) The outermost, youngest wood and the bark of pine. As the sieve cells stop functioning and collapse, the phloem shrinks and becomes undulate. This causes the rays to become wavy. Notice that the phloem rays meet the xylem rays at the ray initials. Also, each row of sieve cells was produced by the same fusiform initial that produced the corresponding row of tracheids (transverse sections; a, X 50; b, X 150).
OUTER BARK
CORK AND THE CORK CAMRIUM
The production and differentiation of secondary xylem cells cause the vascular cambium and secondary phloem to be pushed outward. As the youngest, innermost phloem cells form and mature, they contribute to the larger diameter of the stem or root and increased pressures acting on the outermost tissues. This requires that the tissues on the periphery of the plant either grow in circumference or be torn apart. Actually, the tissues do both, but both must be controlled (Fig. 1). The integrity of the plant surfaces must be maintained against invasion by fungi, bacteria, and insects if the epidermis, cortex, and phloem simply rupture. As circumferential stretching increases and the older sieve elements die, some storage parenchyma cells become reactivated and undergo cell division. This is similar to the activation of parenchyma cells during formation of the interfascicular vascular cambium, but in secondary phloem it results in a new cambium, the cork cambium, also called the phellogen (Fig. 4).
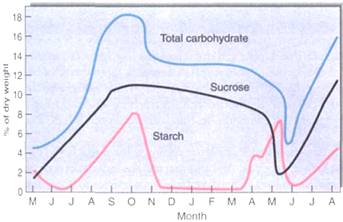
FIGURE 3:Carbohydrate accumulates in the bark of stems and roots during early summer while leaves are still present and photosynthesizing; it reaches its peak in September and October, then drops and remains steady through winter (December to April). It is released during the spring growth season, April and May. These data are for black locusts near Ottawa, Canada, where spring comes late. For plants that grow farther south, the spring release occurs earlier. (Based on data published by D. Simonovitch, C. M. Wilson, and D. R. Briggs).
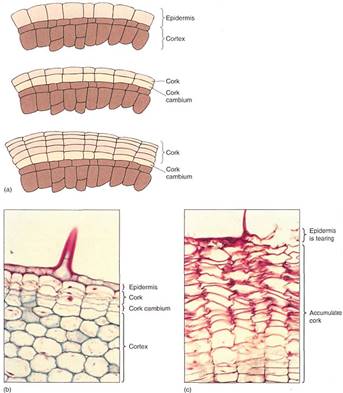
FIGURE 4: (a) The first cork cambium may form as epidermal cells resume mitotic activity. All cell divisions are by periclinal walls, and the inner cell continues as cork cambium while the outer cell differentiates into cork. (b) The hypodermal cells have just started to undergo cell division, resulting in the formation of a cork cambium. This is a young stem of geranium; notice the base of a trichome (X 150). (c) Older stem of geranium; many layers of phellem (cork cells) have accumulated owing to cork cambium activity. Notice that the phellem cells are dead and empty. A trichome was present here also, and the formation of bark blocked transfer of nutrients to the trichome and other epidermal cells, killing them (X 150).
The cork cambium differs greatly from the vascular cambium in both structure and morphogenic activity. All its cells are cuboidal, like ray initials. After each division, the inner cell almost always remains cork cambium while the outer cell differentiates into a cork cell (also called a phellem cell). In a few species, the cork cambium may produce a cell or two to the inside which matures into a layer of parenchyma called phelloderm. The cork cambium, the layers of cork cells, and the phelloderm (if any) are known as periderm. Maturing cork cells increase slightly in volume; then the thin primary walls become encrusted with suberin, making them waterproof and chemically inert; then they die. Cell death is probably a critical part of maturation because after death the protoplasm breaks down, leaving nothing digestible or nutritious for an animal to eat. In many species, some of the cells deposit secondary walls and mature into lignified sclereids; these usually occur in layers that alternate with cork, resulting in a periderm that is both impervious and tough (Fig. 5). Because periderm is such an impermeable barrier, all plant material exterior to it, such as epidermis, cortex, and older secondary phloem, dies for lack of water and nutrients.

FIGURE 5 :(a) A thick layer of uncollapsed cork cells is present on this stem. Many parenchyma cells in the cortex and secondary phloem have converted to sclereids (b), making the bark stronger and more protective. The next cork cambium will form deep in the secondary phloem, where numerous parenchyma cells are capable of becoming mitotically active again; the sclereids cannot resume cell division (transverse sections; a, X 40;b, X 150).
Periderm offers only temporary protection, because the root or stem continues to grow interior to it, pushing it outward and stretching it circumferentially. Unlike vascular cambium, cork cambium is typically short-lived; it produces cells for only a few weeks, after which all cells differentiate into cork cells and die. The layer of cork cells cannot expand much circumferentially, and after one or several years a new cork cambium must be formed in younger secondary phloem closer to the vascular cambium. These new cork cells act as a further barrier and also block water and nutrients from reaching any secondary phloem cells located between layers of cork cambium. In this fashion, several layers of cork can build up.
All tissues outside the innermost cork cambium comprise the outer bark (Fig. 6). All the secondary phloem between the vascular cambium and the innermost cork cambium is the inner bark. The amount of bark is quite variable from species to species; in some only a small amount of cork is formed, so the bark is thin and consists mostly of dead secondary phloem; in others, cork is produced in large amounts and becomes 3 or 4 cm thick. Although it is usually not obvious, bark is continuously falling off the tree, but it does not accumulate at the base because wind and water carry it away.

FIGURE 6: (a) As this pecan trunk increases in diameter, the bark is stretched and ultimately cracks. The deepest bark is the youngest, that on the surface the oldest. (b) The bark of maple peels off in large thin sheets because numerous cork cambia form close together and each is sheetlike. (c) The cork cells of sycamore contain many chemicals; as outer patches of bark peel away, fresh patches are exposed. When their pigments oxidize, they turn grey. Each cork cambium forms as a small patch, the size and shape of the cork cambia affect the nature of the bark. (d) The bark of cork ok (Quoas suber) becomes extremely thick and is composed mostly of phellem, with few sclereids. Cork oaks grown in Spain and Portugal provide most of our commercial corks for baffles. When the bark has become sufficiently thick, the outer bark is peeled away; after a few years, the bark is once again thick enough to harvest.
LENTICELS AND OXYGEN DIFFUSION
The impermeability of cork has negative as well as advantageous consequences. While it keeps out pathogens and retains water, it also blocks absorption of oxygen, interfering with the respiration of the sapwood, vascular cambium, and inner bark. Bark becomes permeable to oxygen when cork cambium produces cork cells that become rounded as they mature. Because rounded cells cannot fit tightly together, intercellular spaces through which oxygen diffuses penetrate the cork layer. These regions of aerenchymatous cork are lenticels (Fig. 7). When a new cork cambium arises interior to this one, it too forms a lenticel in the same place; the outer and inner lenticels are aligned, permitting oxygen to penetrate across all layers of the bark. Lenticel-producing regions are more active than adjacent regions that produce only ordinary impermeable cork; consequently lenticels contain more layers of cells and protrude outward. In species that have smooth bark, even small lenticels are easy to identify. On plants that have thick rough bark, lenticels can be almost impossible to see, but generally they are located at the bases of the cracks in the bark. When the bark of cork oak is made into bottle corks, it is necessary to cut them so that the lenticels do not run from the top of the cork to the bottom.
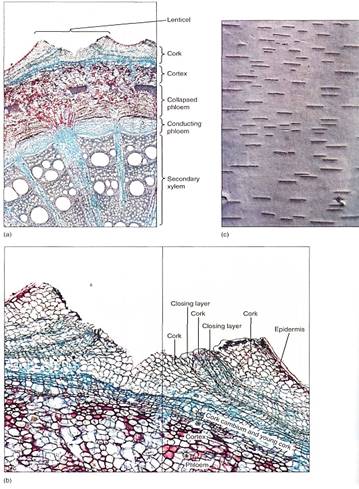
FIGURE 7: (a) Low magnification of stem of Aristolochia, with a large lenticel (X 15). (b) The rapid production of cork cells in the lenticel causes the epidermis to rupture. Although they cannot be seen here, small intercellular spaces allow oxygen to diffuse through the lenticel into the trunk or root (X 50). (c) The lenticels of birch (Betula) become wider as the trunk increases in circumference. Occasionally they divide into two lenticels when the central cork cambium cells begin producing compact cork instead of aerenchymatous cork.
INITIATION OF CORK CAMBIA
The timing of initiation of the first cork cambium is far more variable than that of the vascular cambium. In some species, the first cork cambium arises before a twig or root is even I year old. On stems this is often detectable as the surface color changes from green to tan. In other species, the first cork cambium forms only when that region is several years old, until then, the epidermis and cortex are retained. Epidermises more than 40 years old have been reported. The first cork cambium may arise in a number of tissues: epidermis, cortex, primary phloem, or secondary phloem. Subsequent cork cambia may form shortly afterward, sometimes in the same season, but usually a year or two later. If the growth in diameter is slow, new cork cambia may arise at intervals of as much as 10 years. These later cork cambia usually form deep in the secondary phloem.
The first bark on young stems usually differs from bark formed when the stem is older. If the first cork cambium arises by reactivation of epidermal cells, the first outer bark contains only periderm and cuticle and is very smooth. If the first cork cambium arises in the cortex, the first outer bark contains periderm, cortex, and epidermis; this too is smooth and contains any cortical secretory cells that were present. As the first bark is shed and later cork cambia arise in the secondary phloem, they produce an outer bark that contains only cork and phloem. The nature of these later barks depends greatly on the cell types present in the trapped secondary phloem: Fiber cells produce fibrous, stringy bark, sclereid-filled phloem produces hard bark, and so on. It is not unusual for the bark of a young tree to be dramatically different from the bark it will have when it is older (Fig. 8).

FIGURE 8 :This trunk of Casuarina (red beefwood) still has fragments of its first bark, but the underlying newer bark is also visible. Young branches have only the first type of bark, older parts of the trunk only the second type.
SECONDARY GROWTH IN ROOTS
The roots of gymnosperms and woody dicots undergo secondary growth, as do the stems. A vascular cambium arises just like the interfascicular cambium, when parenchyma cells located between the primary xylem and primary phloem become active mitotically, as do pericycle cells near the protoxylem (Fig. 9). The new vascular cambium has the same star shape as the primary xylem, but it soon becomes round as the cambium in the sinuses of the primary xylem produces more secondary xylem than do the regions of cambium near the arms of protoxylem. Consequently, some portions of the cambium are pushed outward me rapidly than others. When a circular cambium is achieved, the unequal growth stops, and all parts grow at similar rates.
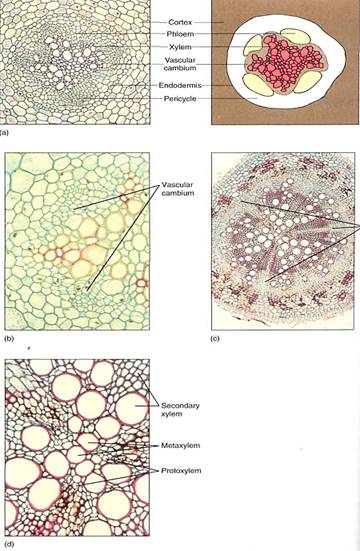
FIGURE 9 :(a and b) Young roots. The vascular cambium of a root such as this baneberry arises as an undulate cylinder located around the xylem, interior to the phloem (a, X 50, b, X 150). (c and d) Older roots. The organization of root wood is similar to that of stem wood: an axial system and a set of rays. Notice in (c) that there are three large rays; the high magnification of (d) reveals that these rays are aligned with the protoxylem of the primary xylem (c, X 40; d, X 150).
The root vascular cambium contains both ray and fusiform initials, and in many cases, wood produced in the root is quite similar to that of the shoot, having sapwood and heartwood and being ring porous or diffuse porous as the stem is. Typically, however, the wood of roots is not identical to that of stems of the same plant, and they may be totally dissimilar when the conductivity requirements of root and stem differ. For example, many cactus roots are extremely long, nonsucculent, cablelike structures. After a brief rain, the numerous root tips absorb water from a large region of surface soil. Because the soil can dry within hours after such a rain, water must be conducted rapidly and in large quantity into the succulent cactus body where it can be stored. Once in the body, however, conduction requirements are totally different; the body may be quite small and consist mostly of parenchyma, with very little wood and just a few small vessels. The water is stored so effectively that most conduction is probably by diffusion through the cytoplasm rather than by the xylem. Furthermore, the narrow roots are strengthened by numerous wood fibers, whereas the small succulent shoots are strengthened only by turgor. In the stem the rays are gigantic, much more voluminous than the axial vessels, providing a large region for water storage. In the roots, which do not store water, rays are small and narrow and consist mostly of sclerenchyma.
Perennial roots also form a bark; the first cork cambium usually arises in the pericycle, causing the endodermis, cortex, and epidermis to be shed. The cork cambium produces cork cells to the outside, forming a protective layer, and in some species a layer or two of phelloderm as well. Lenticels also occur, being especially prominent near lateral roots. The bark on roots may be similar to that on the stem of the same plant, but in some species there are significant differences, again related to the differing metabolisms and microhabitats of the two organs.
Several mechanisms exist by which the storage capacity of a woody root, such as the carrot, can be increased. The ray parenchyma of the secondary xylem offers considerable volume, and storage capacity is increased if the rays become larger. Axial parenchyma in the wood can also be used, and in many storage roots the wood is almost pure parenchyma.



|
|
|
|
4 أسباب تجعلك تضيف الزنجبيل إلى طعامك.. تعرف عليها
|
|
|
|
|
|
|
الفلبين.. بركان "كانلاون" يطلق عمودا من الرماد بارتفاع 4 كم
|
|
|
|
|
|
|
أصواتٌ قرآنية واعدة .. أكثر من 80 برعماً يشارك في المحفل القرآني الرمضاني بالصحن الحيدري الشريف
|
|
|