
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Contorl of Protein Levels |


|
|
Read More
Date: 29-10-2016
Date: 15-11-2016
Date: 18-10-2016
|
Contorl of Protein Levels
As cells undergo differentiation and morphogenesis, their metabolism and structure become different from those of other cells, owing to the presence of proteins, especially enzymes, unique to that cell type. A central question in developmental biology is the mechanism by which distinct cell types control the activities of genes so that they undergo the proper differentiation and obtain the proper set of proteins. There are several points at which protein synthesis and activity theoretically could be controlled: making a gene physically available for transcription; nature of the promoter region; processing of hnRNA into mRNA; transport of mRNA from nucleus to cytoplasm; binding of mRNA to the ribosome small subunit; rate of translation; processing of protein; activation or inactivation of the protein.
Biochemical studies have shown that many enzymes and structural proteins may be present in a cell in an inactive form. Tubulin is an excellent example: A cell may contain a large pool of tubulin monomers, then aggregate them into microtubules at a specific lime Microtubules can appear rapidly without the need for gene activation or protein synthesis. Similarly, enzymes are often completely inactive until phosphate groups are added by a class of enzymes, called kinases, that are themselves activated by the arrival of hormones. Hormones arrive at the plasma membrane and bind to receptors, which then synthesize second messengers that enter the cell. These activate the phosphorylating enzymes that in turn activate the dormant enzymes, leading to a significant change in the cell's metabolism. This is mostly an activating rather than a differentiating mechanism; these cells are already prepared for highly specific responses.
It would be extremely inefficient if a cell transcribed all its genes and synthesized all of its possible proteins when only a lew are needed. For maximum efficiency, a cell should not even synthesize the mRNA for proteins it does not need. We expect that the most fundamental level of control of morphogenesis would occur at the level of transcription.
Unfortunately, we know very little about the control of transcription in eukaryotas, especially plants. Our lack of specific knowledge in this area is not due to lack of effort but to the tremendous technical problems involved. Except for the genes that code for rRNA and tRNA, most genes are present as only two copies in a diploid nucleus. When hormones cause a cell to produce a new protein, it may be that only one or two activator molecules bind to the promoter region of one or both copies of the gene that codes for that protein. Consequently, when we study the activation of a gene, we study a process in which a change occurs in only one or two molecules out of the billions in the nucleus. Because genes can also be turned off, the chemical messengers can also release from DNA, making it extremely difficult to find the actual molecules responsible for activating a particular gene.
Theoretically, chemical signals could activate genes in several ways. The hormone/receptor complex might actually bind to DNA, or presence of the hormone might cause the receptor to be liberated from a membrane so that the receptor alone binds to DNA. Or the hormone/receptor complex might cause synthesis or release of a second messenger that then enters the nucleus. Although past research in this area has been slow, DNA analysis techniques offer great promise. The genes for many proteins have now been identified, and their promoter regions are being studied. It is hoped that genes turned on simultaneously have aspects of their promoters that will indicate how they are activated in unison. For example, phenolic compounds are important in many aspects of a plant's metabolism, and their biosynthetic pathway is well understood. When cell cultures of parsley, which have few phenolics, are transferred from darkness to light, phenolic pigments called flavonoids appear. This is preceded by the sudden synthesis of the first three enzymes of their metabolic pathway, followed a few hours later by the appearance of the next two enzymes (Fig.). The first three enzymes may be coded by genes whose promoters are identical or are activated by similar chemical messengers.
Control can be exerted in the processing of hnRNA into mRNA. Rather surprisingly, the great majority of RNA transcribed in nuclei is immediately degraded back to free nbonucleotides. Even accounting for the large number of introns that must be removed, many exons are probably degraded also. Even though a gene is transcribed, its information may not be converted into mRNA. Some of the chemical messengers from the cytoplasm my bind to hnRNA rather than to DNA; they would regulate hnRNA processing rather than DNA transcription.
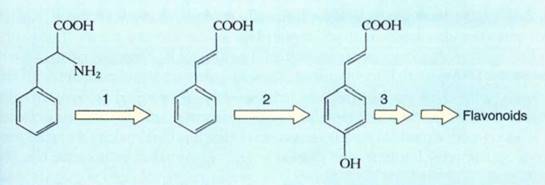
FIGURE :The genes for the first three enzymes in this pathway for the synthesis of flavonoids and lignin are turned on simultaneously. Perhaps they have identical promoters and are activated by the same type of intracellular chemical messenger. The enzymes are (1) phenylalanine ammonia lyase, (2) cinnamate 4-hydroxylase, and (3) 4-coumarate: CoA ligase.



|
|
|
|
دراسة يابانية لتقليل مخاطر أمراض المواليد منخفضي الوزن
|
|
|
|
|
|
|
اكتشاف أكبر مرجان في العالم قبالة سواحل جزر سليمان
|
|
|
|
|
|
|
اتحاد كليات الطب الملكية البريطانية يشيد بالمستوى العلمي لطلبة جامعة العميد وبيئتها التعليمية
|
|
|
















