























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Vascular Cambium
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 15-11-2016
15-11-2016
 6270
6270




+
-
20
Vascular Cambium
INITIATION OF THE VASCULAR CAMBIUM
The vascular cambium (plural: cambia) is one of the meristems that produce the secondary plant body (Fig.1). In an herbaceous species, the cells located between the metaxylem and metaphloem of a vascular bundle ultimately stop dividing and differentiate into conducting tissues.

FIGURE 1: (a) The cambium has recently formed in this Bocconia stem. The fascicular cambia are rather obvious, but the interfascicular cambia are difficult to see. They have produced more secondary xylem than secondary phloem (X 180). (b and c) This cambium has only recently iormed; it contains two or three layers of secondary xylem and one or two of secondary phloem. Three croups of primary xylem are visible, so you can tell where fascicular and interfascicular cambia arose (b, X 50; c, X 250).
But in a woody species, the cells located in this position never undergo cell cycle arrest; they continue to divide instead of maturing, and they constitute the fascicular cambium (Fig. 2a). In addition, some of the mature parenchyma cells between vascular bundles come out of cell cycle arrest and resume mitosis, forming an interfascicular cambium that connects on each side with the fascicular cambia. Once this happens, the vascular cambium is a complete cylinder. The terms "fascicular" and "interfascicular" are used only while the cambium is young; after 2 or 3 years the two regions are usually indistinguishable, and then only the term "vascular cambium" is used.
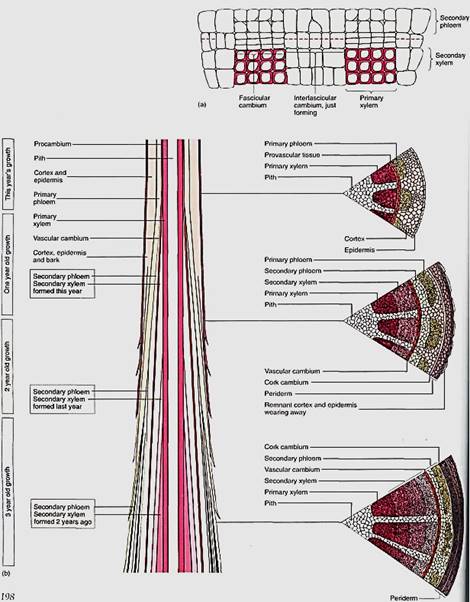
FIGURE2: (a) Two vascular bundles and the parenchyma located between them. Some of the parenchyma cells have begun renewed cell division and constitute an interfascicular cambium. This zone of renewed mitotic activity is located between two fascicular cambia, so one full cambial zone will result. (b) The vascular cambium usually does not form until after a portion of shoot or root is several weeks or even many months old, so branch tips have only primary growth. Once a region is old enough, the vascular cambium forms and secondary xylem begins to accumulate to the inside of the cambium; secondary phloem accumulates to the exterior. At first, the vascular bundles are still recognizable. After several years of activity, considerable amounts of secondary xylem and phloem accumulate.
Vascular cambia must be extended each year. The tips of roots and stems initially contain only primary tissues, but at some time in the same season, a vascular cambium arises and that portion of the root or stem then contains both primary and secondary tissues (Fig. 8.3b). During the next growing season, the apical meristem extends the axis beyond this point; a new segment of vascular cambium forms within it and joins at its base to the top of the vascular cambium formed in the previous season. The vascular cambium within a tree consists of segments of distinct ages, those near the ground being oldest and those closer to the tips of the axes being younger.
Very rarely a vascular cambium forms in leaves that stay on a tree for many years, but just a tiny amount of secondary tissues, usually only secondary phloem without any secondary xylem, is formed in the midrib; the other veins contain only primary tissues. Secondary growth never occurs in flowers, fruits (except some fruit peels that are actually bark), or seeds.
Although a vascular cambium shares many features with an apical meristem, it is unique in certain aspects. It is a rather simple meristem in that it has only two types of cells, fusiform initials and ray initials (Figs. 3 and 4).
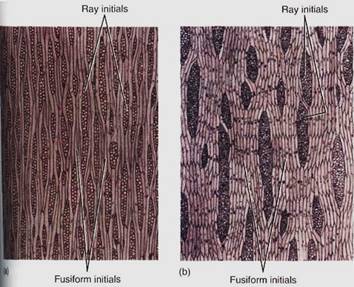
FIGURE 3:Tangential sections through a nonstoried vascular cambium of apple (Malus sylvestris, a) and a storied vascular cambium of black locust (Robinia pseudoacacia, b). Notice that the storied fusiform initials are extremely long and the ends are not aligned. In the storied cambium, the fusiform initials are much shorter and occur in horizontal rows. (Courtesy of R. Evert, University of Wisconsin)
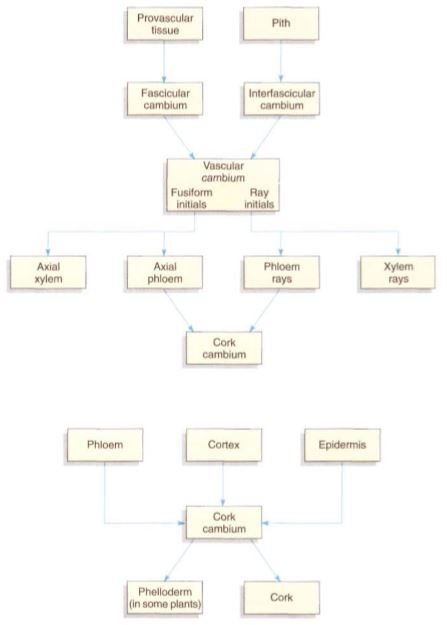
FIGURE 4: Cell lineages during secondary growth.
FUSIFORM INITIALS
Fusiform initials are long, tapered cells; typical lengths for fusiform initials are 140 to 462 µm in dicots and 700 to 8700 µm in gymnosperms. When a fusiform initial undergoes longitudinal cell division with a wall parallel to the circumference of the cambium (a periclinal wall), it produces two elongate cells (Fig. 5). One continues to be a Mom initial, and the other differentiates into a cell of secondary xylem or secondary phloem. If the outer daughter cell remains a cambium cell, the inner cell develops into secondary xylem. But if the inner one continues as cambium, the outer cell differentiates and matures into secondary phloem.. This orientation is constant: Wood never forms to the exterior of the vascular cambium, and bark never forms on the interior side. Regardless of which cell differentiates, one always remains as cambium. It is not known what factors determine which cell remains cambial and which differentiates, but within any year, both xylem and phloem are produced, almost always much more xylem than phloem.
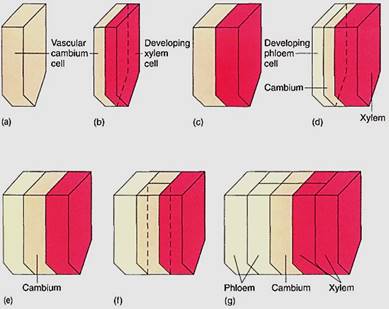
FIGURE 5: (a) The lower half of a fusiform initial before division (to simplify the diagram, the top half has not been drawn in). (b) Division by a periclinal wall results in two thin cells; the outer one remains a fusiform initial and the inner cell develops into secondary xylem. (c) Both cells enlarge to the size of the original cell. (d) The fusiform initial divides again; this time the outer cell matures as secondary phloem while the inner one remains a fusiform initial. (e) The cells grow back to the original size. (f) The fusiform initial divides by an anticlinal wall, resulting in two fusiform initials. (g) After the radial division in (f), a new row of cells is initiated in the secondary xylem and phloem.
Cambial cells produce narrow daughter cells, all of which enlarge during differentiation. Daughter cells located on the inner side, which mature into secondary xylem, increase greatly in diameter, causing the cambial cells to be pushed outward (Figs. 6 and 7).
Because the cambium is a cylinder, such outward movement results in a cylinder of larger circumference. Vascular cambium cells must divide longitudinally by anticlinal walls, Thereby increasing the number of cambial cells (Fig. 8.6f). Without anticlinal divisions, cambial cells would be stretched wider circumferentially and finally could not function .
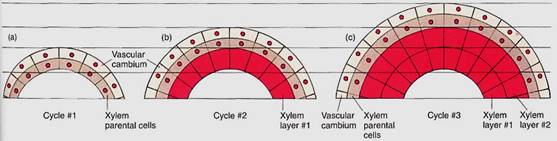
FIGURE 6: Afte vascular cambium cells divide, the progeny cells on the interior side become xylem parental cells. As all cells expand to their mature size, the cambial cells are pushed outward. The cambial cells divide again, depositing a new layer of xylem parental cells exterior to the xylem that just fomad (xylem layer 1). The new xylem parental cells expand, pushing the vascular cambium farther outward. Cambial cells divide again. Notice that each xylem cell is formed in place; they are not pushed inward.
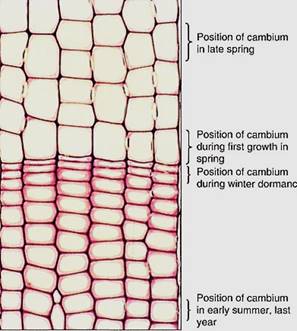
FIGURE 7: Six rows of wood cells are visible in this transverse section of a pine trunk. All the cells of each row were produced by a single vascular cambium cell, so all the wood visible in this micrograph was produced by just six cambium cells. The cells at the bottom of the picture were produced when the cambium cells were located at that site; similarly, the cells in the middle of the picture were produced when the cambium exited there, having been pushed outward by the production of the cells at the bottom of the picture (X 150).
RAY INITIALS
Ray initials are similar to fusiform initials except that they are short and more or less cuboidal. They too undergo periclinal cell divisions, with one of the daughters remaining a cambial ray initial and the other differentiating into either xylem parenchyma if it is the inner cell, or phloem parenchyma if it is the outer cell. One of the most significant differences between fusiform and ray initials is that the elongate fusiform initials produce the elongate cells of the wood (tracheids, vessel elements, fibers) and the phloem (sieve cells, sieve tube members, companion cells, fibers). Ray initials produce short cells, mostly just storage parenchyma and, in gymnosperms, albuminous cells.
ARRANGEMENT OF CAMBIAL CELLS
Ray and fusiform initials are organized in specific patterns. Ray initials are typically grouped together in short vertical rows only one cell wide (a uniseriate ray; see Fig. 3), two cells wide (a biseriate ray), or many cells wide (multiseriate). Fusiform initials may occur in regular horizontal rows (a storied cambium) or irregularly, without any horizontal pattern (a nonstoried cambium). Storied cambia have envolved more recently than nonstoried cambia and occur in only a few advanced dicot species, for example, redbud and persimmon. The fusiform initials of storied cambia tend to be short, less than 200 pm long. The selective advantage of storied tissues is unknown.
Typically, a vascular cambium never has large regions of just fusiform initials or just ray initials. If anticlinal divisions result in many fusiform initials side by side, a central one may undergo transverse divisions and be transformed into a set of ray initials . Likewise, if a group of ray initials becomes unusually broad, one or several central ones may elongate and be converted to a fusiform initial. The ratio of fusiform initials to ray initials within a species is quite constant and apparently under precise developmental control.
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















