
آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Anomalous Forms of Growth |


|
|
Read More
Date: 23-10-2016
Date: 2-11-2015
Date: 28-10-2015
|
Anomalous Forms of Growth
ANOMALOUS SECONDARY GROWTH
The development, cellular arrangement, and activity of the vascular cambia in gymno- sperms and most woody dicots are remarkably similar. There are alternatives to this type of vascular cambium, and an analysis of the consequences of various types of arrangements or activities can reveal a great deal about secondary growth. Because these cambia produce secondary bodies that differ from the common type, their growth is called anomalous secondary growth.
Roots of Sweet Potatoes. In sweet potatoes (Ipomoea batatas), the amount of storage parenchyma is increased dramatically by an anomalous method of secondary growth.
Numerous vascular cambia arise, not around the entire mass of primary xylem, but around individual vessels or groups of vessels (Fig. 1). The cambia act normally, except that the xylem and phloem produced are almost purely parenchyma. New vessels may also be surrounded by another new cambium, and the process is repeated. As the sweet potato becomes quite large, it may contain hundreds of cambia of various ages; the secondary tissues are an irregular matrix of parenchyma, a few sieve tubes, some vessels, and vascular cambia.
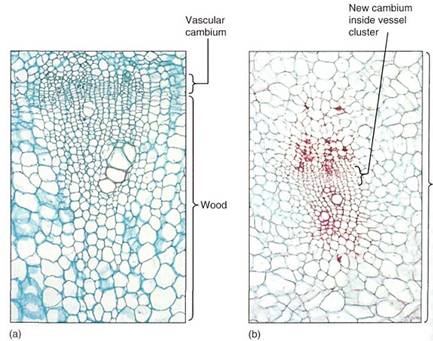
FIGURE 1: (a) The part of a radish that we eat is the wood of the storage root. It does not look like wood at first glance. The wood is mostly parenchyma with a few vessels and no fibers. This anomalous wood is well-adapted for the rapid production of, and conduction into, long-term storage tissue (X 40). (b) In the wood of beets (again, the part that we eat), additional cambia arise within clusters of vessels (X 40).
What is the selective advantage of producing so many vascular cambia instead of just one? It may be that rate of cell production is important. Because the root must become large very quickly, having just one vascular cambium may be too slow. Multiple vascular cambia all functioning simultaneously speed up the production of storage capacity.
Included Phloem. In several dicots, a vascular cambium of the common type arises and produces ordinary secondary xylem and phloem. But after a short period, the cambium cells stop dividing and differentiate into xylem, and there is no cambium. But cells in the outermost, oldest secondary phloem become reactivated and differentiate into a new vascular cambium that acts just like the first, producing ordinary secondary xylem and phloem, then differentiating and ceasing to exist. Notice in Figure 8.30 that because the second cambium arose in the outermost phloem, the xylem it produces is located exterior to the phloem of the first cambium. From interior to exterior there is first xylem, first phloem, second xylem, second phloem. Then a new vascular cambium arises in the outermost second phloem, and so on. This type of secondary phloem, located between two bark of xylem, is included phloem. The selective advantage may be protection of the phloem from insects and other pests by one to several layers of wood. Also, the tissue relationships between xylem and included phloem differ from those between xylem and ordinary phloem, but the advantages of this have not yet been studied.
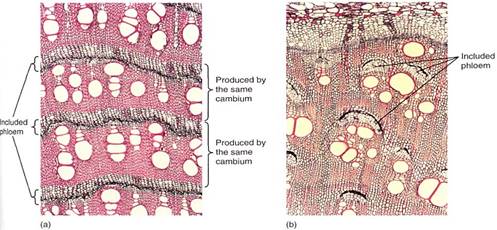
FIGURE 2 :(a) This stem of Iresine shows bands of secondary xylem alternating with bands of included phloem. Each band of xylem and the phloem just external to it were produced by one cambium; the next set was produced by a different cambium (X 40). (b) Bougainvillea also has included phloem, but it occurs in patches, not complete bands (X 40).
Unequal Activity of the Vascular Cambium. In ordinary growth, all areas of the vascular cambium have about equal activity, so the stem or root is round in cross-section. But in some species of Bauhinia and certain other woody vines, two sectors of the cambium are very active while two are almost completely inactive (Fig. 3), so the stem grows outward in two directions but remains thin in the other two and soon becomes a thin, flat, woody ribbon. The selective advantage of this may be related to the relationship between conductivity and flexibility. As an ordinary round stem becomes wider in all directions, its conducting capacity increases but its flexibility decreases. As the Bauhinia stem becomes wider, its conducting capacity increases but its flexibility remains about the same. For many vines, flexibility is selectively advantageous, and this type of anomalous secondary growth is adaptive.
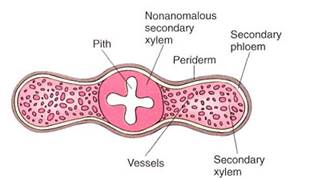
FIGURE 3:The cambium on two opposite sides of this Bauhinia is rapidly producing cells, many of which differentiate into wide vessel elements. The two alternate portions of cambium produce very few cells, and all of them mature without much expansion.
Secondary Growth in Monocots. None of the monocots has secondary growth like thatin dicots, but some do become treelike and "woody," such as Joshua trees, dragon trees, and palms. The first two groups undergo a process of anomalous secondary growth; the palms have an unusual type of primary growth.
In Joshua trees (some members of the genus Yucca; Fig. 4) and dragon trees (in the genus Dracaena), a type of vascular cambium arises just outside the outermost vascular bundles (Fig. 5) which originates from cortex cells in the same manner that the interfascicular vascular cambium arises in dicots. This cambium, however, produces only parenchyma; conducting cells are completely absent. Some of the parenchyma cells undergo rapid division and produce columns of narrow cells that differentiate into secondary vascular bundles containing xylem and phloem. The outermost cells of each bundle develop into fibers with thick secondary walls. The parenchyma cells that do not divide like this form a secondary ground tissue, the arrangement of which is almost identical to that of primary tissues. They are "woody" because of the fibers, and they have more conducting capacity and greater strength each year, so branching is feasible.

FIGURE 4: Joshua tree (Yucca brevifolia) is an arborescent monocot that belongs to the lily family. Because it has secondary growth, even though of an unusual type, its ability to conduct increases, and both branching and increased numbers of leaves are feasible without adventitious roots of the type necessary for screwpine (see Fig. 7.18).
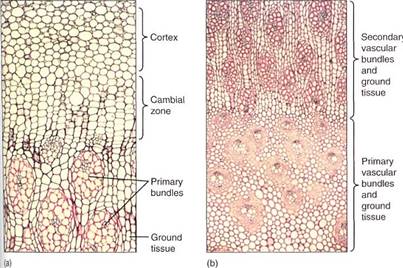
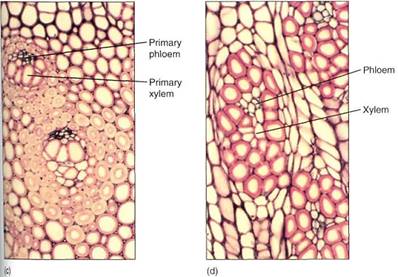
FIGURE 5:Seocndaiy growth in arborescent monocots. (a) The broad zone of the vascular cambium region and the immature secondary ground tissues are visible. The bundles are secondary, and a secondary cortex also forms (X 50). (b) The inner part of the stem, with the primary vascular birdies and ground tissue on the bottom and the secondary bundles at the top (X 15). (c) Primary bundles (X 50). (d) Secondary bundles (X 50).
A palm seedling does not have a full set of leaves and a wide trunk. For the first few years of life, the palm trunk becomes wider and the number of leaves increases. This happens without secondary growth because during the seedling years, palms produce numerous adventitious roots from the base of the short stem. Each root adds extra vascular bundles, and the portion of stem above each new root can have that many more bundles than it does below the root. For example, if the stem has 100 bundles at one point and if just above this it produces five adventitious roots that each have eight bundles, then above these roots the stem can have 140 bundles. This increase in width and addition of adventitious roots in palms is establishment growth, a form of primary growth. At some point this process ceases, no new adventitious roots are established, and the conducting capacity is set for the lifetime of the plant. This same type of primary growth occurs in other monocots with extremely broad stems such as corms and bulbs.



|
|
|
|
دراسة يابانية لتقليل مخاطر أمراض المواليد منخفضي الوزن
|
|
|
|
|
|
|
اكتشاف أكبر مرجان في العالم قبالة سواحل جزر سليمان
|
|
|
|
|
|
|
اتحاد كليات الطب الملكية البريطانية يشيد بالمستوى العلمي لطلبة جامعة العميد وبيئتها التعليمية
|
|
|
















