آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Flowering |


|
|
Read More
Date: 17-10-2016
Date: 27-10-2016
Date: 31-10-2016
|
Flowering
RIPENESS TO FLOWER
Almost all plants must reach a certain age before they can be induced to flower; only a species can be induced as seedlings. Annual plants need to be only several weeks old before they become competent to respond to a floral stimulus, but many perennials must be 5, 10, or even 20 years old. Before this time, conditions that should induce flowering have no effect. Virtually nothing is known about the metabolic difference between the juvenile stage, when plants are incapable of being induced to flower, and the adult stage, when they are sensitive to floral stimuli.
Cold temperatures are the stimulus responsible for converting biennial plants to the adult condition, in the process of vernalization. The site of perception is the shoot apex itself; if it is cooled while the rest of the plant remains warm, vernalization occurs. But if the rest of the plant is cooled while a small heater keeps the apex warm, no vernalization occurs. Presentation time is as short as 1 day in some plants. The transduction process is known to require oxygen, carbohydrates as an energy source, ad an optimal temperature just above freezing, between 1 and 7°C. Vernalization results in a stable change. If the plants are returned to warm conditions but are not given the fled stimulus of short days and long nights, they continue to grow vegetatively, without flowering, year after year. But they retain their vernalization and flower whenever the floral stimulus is finally given.
PHOTOPERIODIC INDUCTION TO FLOWER
The conversion of an adult plant from the vegetative to the flowering condition may be the most complex of all morphogenic processes. This is not one process—different mechanisms exist in different species. In certain annual species, size appears to be the only important factor: Peas and corn initiate flowers automatically after a particular number of leaves has been produced, regardless of environmental conditions; flowering is controlled by internal mechanisms. In many species, perhaps most, transition to the flowering condition is triggered by photoperiod—day length—which acts as a season indicator (Table 1). One subclass of these plants blooms when days are short (spring or fall) and are short-day plants. Another subclass, long-day plants, are induced to bloom when days are long, in summer. Plants that do not respond to day length are day-neutral plants.
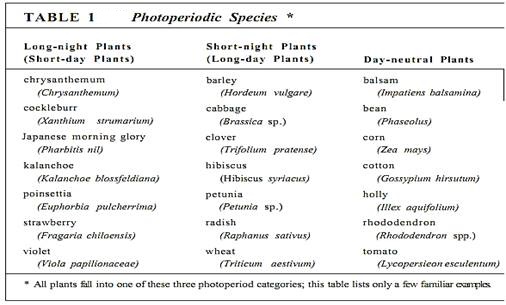
We know much about how plants measure day length. First, the pigment phytochrome detects the presence or absence of light (Table 2; Fig. 1). Phytochrome has a light-absorbing portion attached to a small protein of about 124,000 daltons. When phytochrome absorbs red light with a wavelength of about 660 nm, the protein changes its folding. This affects many of its properties, one of which is its hydrophobicity; in the refolded state, it is more hydrophobic and binds to membranes. A second altered property is its absorption spectrum; it now absorbs not at 660 nm but in the far-red (almost infrared) region of 730 nm. But when this form absorbs far-red light, it refolds back to the red-absorbing form and releases from the membrane. The two forms are called Pr (red-absorbing) and Pfr (far red-absorbing). Also, Pfr reverts to Pr in darkness. Apparently Pr is inactive metabolically but becomes morphogenically active and exerts its effect when it absorbs red light and is converted to Pfr: Pfr is the active form and may bring about metabolic responses. Air a plant is given red light, converting phytochrome to the active Pfr form, exposure to fined light converts the phytochrome back to the inactive Pr form. If far-red light is given quickly enough after red light, phytochrome does not have enough time to affect cell metabolism significantly, and no effect is seen. But if the far-red comes long enough alter the red for the presentation time to be fulfilled, the Pfr is able to complete the transduction process and far-red light can no longer cancel the red light stimulation.

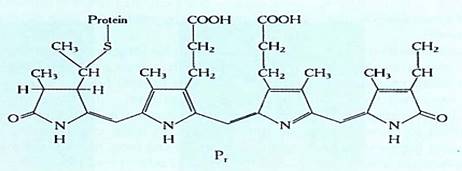
FIGURE 1:The chromophore or light-detecting portion of phytochrome. The rest of the molecule is a protein, Notice that the two portions are joined by a sulfur atom.
A plant experiencing a short day in nature automatically receives a long night (Fig. 2). Similarly, long days are always accompanied by short nights. Night length is actually the critical factor. A long-day plant is in reality a short-night plant. It can be placed in a growth chamber and artificially given both long days and long nights—for example, 16 hours of light and 16 hours of dark in a 32-hour "day." If day length is the important factor, the plant should flower because it has long days; if night is the critical factor, then it should not flower because it does not have short nights. When the experiment is done, the long-day plant does not flower, indicating that night length is critical, not day length. Similar experiments have shown that short-day plants really are long-night plants: If given a 16-hour cycle (8 hours light/8 hours dark—both day and night are short), they do not flower. But if given long nights, even accompanied by artificially long days, the plants are induced to flower.

FIGURE 2 :(a) and (b) Natural 24-hour light/dark cycles must have short nights any time the days are long (summer) and long nights when days are short (winter), so plants could detect season by measuring either day or night. When photoperiodism was first discovered, it was assumed that day length was important, so all our terminology was based on that. (c) With electric lights, it is possible to create a long photoperiod—36 hours long here—with both long days and long nights. Under these conditions, short-day (long-night) plants bloom, but long-day (short-night) plants do not.
Each species has its own particular requirements for long or short nights; that is, not all "long nights" have to be the same length. Instead each species has a critical night length; if a short-night plant receives nights shorter than this critical length, it flowers, whereas a long-night plant must receive nights longer than its own critical night length. Baas critical night length varies from species to species, it is possible for a long-night plant and a short-night plant to bloom under the same conditions if the critical night length for the short-night plant happens to be longer than that for the long-night plant (Table 3).

Day length/night length control of flowering and other processes such as the initiation or breaking of dormancy is more common at locations farther from the equator. Away from the equator, nights become progressively shorter from winter to summer, then progressively longer from summer to winter. The greater the distance from the equator, the greater the length of the longest winter night and the shorter the length of the shortest summer night. Thus, if two species are to bloom just after the beginning of May, a species in the southern United States or Mexico must have a critical night length shorter than that of a species in the northern United States or Canada.
Presentation time varies considerably; in some morning glories, one photoperiod of the proper length induces flowering, whereas at least 1 or 2 weeks of proper photoperiods are necessary for other species. The accuracy with which night lengths can be measured varies, but the most accurate species known is the long-day (short-night) plant henbane (Hyoscyamus niger); it must have nights shorter than 13 hours, 40 minutes. If the nights are even 20 minutes too long, 14 hours long for example, it does not flower.
When phytochrome was discovered to be responsible for measuring night length, it was hypothesized that most phytochrome was converted to Pfr by the end of a day and then reverted slowly back to Pr during the night. It was postulated that if it could completely revert in the dark, before the next sunrise, metabolic changes would be triggered. However, it is now known that virtually all Pfr converts back to Pr within 3 or 4 hours, a time far too short to be a night-measuring clock by itself, although it may be the initial part of a longer-acting clock.
The sites of perception for night length are young leaves. It rs possible to stimulate one leaf with a spotlight and induce the plant to flower, even if the apical meristem, the site of response, is not illuminated (Fig. 3). Because the site of perception is not the site of response, a chemical messenger must be transmitted between the two. If a leaf is photoinduced and then immediately cut off the plant, no flowering occurs; if it is allowed to remain attached for several hours, the flowering stimulus is synthesized and transported out of the leaf. If the leaf is then removed, the plant still flowers. An obvious experiment is to induce a leaf by giving it the proper night length, then collect the sap that is transported through the petiole and assay it for the hormone that acts on the apical meristem. This has been done hundreds of times by many people, without any repeatable success; the process is not as simple as we had at first thought.

FIGURE 3:If a short-night plant is given long nights, it does not flower. But it is possible to cause flowering by illumination with 15 minutes of dim red light; the plant acts as though it has received two short nights separated by a 15-minute day. (a) The red light "night break" does not have to be given to the whole plant; if a narrow beam of red light shines on a single leaf while all the rest of the plant remains in darkness, the plant flowers (b).
An extremely interesting set of results has been obtained by grafting together plants with different photoperiod requirements. Individuals of the tobacco species Nicotiana silvestris are long-day plants. The species N. tabacum has two types of individuals: Those of the cultivar N. tabacum cv. Trabezond are day-neutral plants, and those of N. tabacum cv. Maryland Mammoth are short-day plants. When long-day and day-neutral plants are grafted together and given long days, the day-neutral plant is also induced to flower, presumably by a floral stimulus that passes through the graft union. Similarly, day-neutral plants are induced to flower if grafted to short-day plants and given short days (Fig. 4). Grafting one day-neutral plant to another does not increase flowering, so the grafting by itself has no effect.
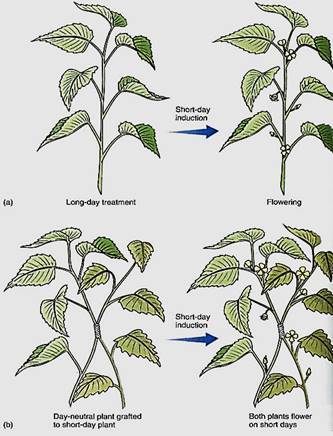
FIGURE 4 :(a) Under short days, short-day plants are induced to flower but day-neutral ones are not. (b) If grafted together and given short days, both short-day plants and day-neutral plants flower. Apparently the floral stimulus that was induced in the short-day plant has been transported to, and is active in, the day-neutral plant.
When the combination of short-day plants grafted to day-neutral plants was given long days, the short-day plants did not flower, as expected, and the day-neutral partners flowered about the time they would have if not grafted to anything. But when long-day plants grafted to day-neutral plants were exposed to non-inductive short days, not only did the long-day plants not flower, but the day-neutral partners were also prevented from flowering. Apparently, under non-inductive conditions, long-day plants actually produce an inhibitor of flowering. Flowering in these long-day tobacco plants seems to be controlled by a switch from inhibitor production to promoter production, whereas in the short-day tobacco plants, flowering is controlled only by the presence or absence of a promoter. Neither the promoter nor the inhibitor has been isolated, but gibberellins are suspected to be involved in the synthesis or activation of the promoter.
ENDOGENOUS RHYTHMS AND FLOWERING
Plants contain endogenous rhythms; that is, certain aspects of their metabolism cycle repeatedly between two states, and the cycle is controlled by internal factors. The most obvious example of this is in the "sleep movements" of the leaves of plants like prayer plant (Oxalis). In the evening, leaflets drop down, and in the morning, they raise themselves to the horizontal position as motor cells increase their turgor. It is easy to assume that this is a photonastic response, but if the plants are placed in continuous darkness, the leaflet position continues to change, returning to the up position about every 24 hours . In many flowers, the production of nectar and fragrance is also controlled by an endogenous rhythm and occurs periodically even in uniform, extended dark conditions.
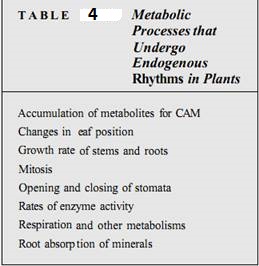
Endogenous rhythms are involved in numerous aspects of plant metabolism that are rot easily observed, such as many aspects of photosynthesis, respiration, growth rate, and exudation of mucilage from roots (Table 4). The underlying mechanism that constitutes the clock is poorly understood but is known to be independent of temperature and general health of the plants. Endogenous rhythms are not affected by cooling or warming. Also, extensive studies have shown that the rhythm is truly endogenous and not controlled by an unsuspected exogenous rhythm related to Earth's rotation. Plants have been taken to the South Pole, where planetary rotation would have no effect, and have been taken into orbit (Fig 5). If the rhythms were actually exogenous, plants at the South Pole should lose their rhythmicity, whereas those in orbit should have a more rapid rhythm that matches the orbital period. In both cases, normal rhythm was maintained.

FIGURE 5 :Ruling out exogenous factors such as light and temperature fluctuations does not prove that rhythms are endogenous; the plants could be responding to an exogenous rhythm we have not yet thought to test. But if it is exogenous, then in orbit the rhythm should match the period of the orbit. Experiments have shown that even in orbit, the 24- hour cycle is maintained. The space shuttle has made such experiments possible. This is F. Chang-Diaz performing an experiment on root growth for Dr. Randy Moore. (Courtesy of NASA, R. Moore, and E. McClellan)
Many types of endogenous rhythms have a period that is not 24 hours long. Cytoplasmic streaming and the spiralling motion of elongating stem tips have periods of only a few minutes to a few hours; these are ultradian rhythms. If a period is approximately 24 hours long, it is a circadian rhythm, the most common kind. The release of gametes in brown algae is controlled by a 28-day lunar rhythm. Some seeds have an annual rhythm of germinability: If stored in uniform conditions and periodically provided with moisture and warmth, they germinate only at times of the rhythm that correspond to springtime.
"Circadian" means that the rhythm is only approximately, not exactly, 24 hours long. When placed in uniform conditions, the true cycle typically differs slightly from 24 hours, being either somewhat longer or shorter (Fig. 6). However, in nature, the rhythm is exactly 24 hours long because light is able to entrain (reset) the rhythm. The pigment responsible for detecting the light for entrainment is phytochrome. Each morning, sunrise resets the rhythm so it can never get out of synchronization with exogenous light/dark cycles.
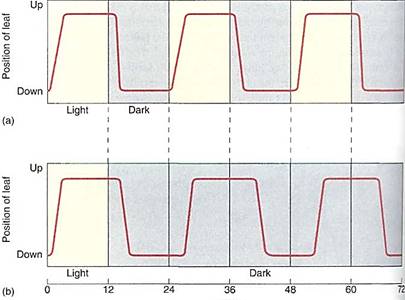
FIGURE 6: (a) Under natural conditions, a circadian rhythm matches the cycle of light and dark, being actually 24 hours long. (b) In continuous darkness, most circadian rhythms have periods slightly longer than 24 hours. In nature, sunrise resets the clock by acting on phytochrome every morning.
The involvement of endogenous circadian rhythms in flowering was discovered during dark interruption experiments: A short-day (long-night) plant can be prevented from flowering by interrupting long nights with a brief (15 minutes or less) exposure of red light:
This is detected by phytochrome, and the plant acts as though it has received two short nights separated by a 15-minute day. Short-day plants given a very long night— continuous darkness—have an endogenous rhythm of sensitivity to light breaks. If the light break is given at 6 hours into the dark period, or at 30 hours (24 + 6), 54 hours (24 + 24 + 6), and so on, the light break prevents flowering (Fig. 7). These times correspond to darkness in a normal environment. But if the light break is given at a time when the endogenous rhythm would be "expecting" normal daylight conditions, such as at 16, 40 (24 + 16), or 64 (24 4- 24 4- 16) hours after the beginning of the dark treatment, the light break does not stop flowering. Plants kept in uniform, dark conditions undergo an endogenous cyclic sensitivity and insensitivity to red light interruption of the critical night length. Just how the endogenous rhythm and the critical night length work together to stimulate flowering is not known.
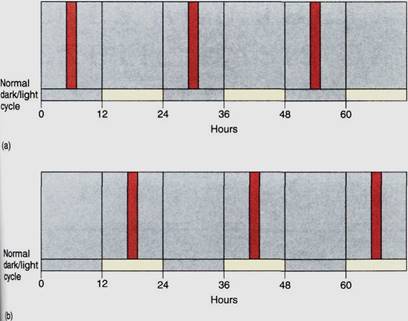
FIGURE 7 :If a long-night plant is placed in continuous darkness, it can be prevented from flowering by giving it red light night breaks, but these are effective only if given at those times of the endogenous rhythm when the plant is "expecting" darkness (a). If given when the rhythm is "expecting" light, the light breaks are ineffective (b). Whatever metabolism red light interrupts does not occur continuously in prolonged darkness, but rather periodically, controlled by the internal clock .



|
|
|
|
التوتر والسرطان.. علماء يحذرون من "صلة خطيرة"
|
|
|
|
|
|
|
مرآة السيارة: مدى دقة عكسها للصورة الصحيحة
|
|
|
|
|
|
|
نحو شراكة وطنية متكاملة.. الأمين العام للعتبة الحسينية يبحث مع وكيل وزارة الخارجية آفاق التعاون المؤسسي
|
|
|