آخر المواضيع المضافة

 النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية
النبات
الحيوان
الأحياء المجهرية
علم الأمراض
التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحياتية النانوية
علم الأجنة
الأحياء الجزيئي
علم وظائف الأعضاء
الغدد
المضادات الحيوية| Liver : Carbohydrate metabolism |


|
|
Read More
Date: 8-12-2021
Date: 21-12-2021
Date: 24-11-2021
|
Liver : Carbohydrate metabolism
The liver is uniquely situated to process and distribute dietary nutrients because the venous drainage of the gut and pancreas passes through the hepatic portal vein before entry into the general circulation. Thus, after a meal, the liver is bathed in blood containing absorbed nutrients and elevated levels of insulin secreted by the pancreas. During the absorptive period, the liver takes up carbohydrates, lipids, and most amino acids. These nutrients are then metabolized, stored, or routed to other tissues. In this way, the liver smooths out potentially broad fluctuations in the availability of nutrients for the peripheral tissues.
Carbohydrate metabolism
The liver is normally a glucose-producing rather than a glucose-using organ. However, after a meal containing carbohydrate, the liver becomes a net consumer, retaining roughly 60 g of every 100 g of glucose presented by the portal system. This increased use reflects increased glucose uptake by the hepatocytes. Their insulin-independent glucose transporter (GLUT-2) has a low affinity (high Km [Michaelis constant]) for glucose and, therefore, takes up glucose only when blood glucose is high . Processes that are upregulated when hepatic glucose is increased include the following.
1. Increased glucose phosphorylation: The elevated levels of glucose within the hepatocyte (as a result of elevated extracellular levels) allow glucokinase to phosphorylate glucose to glucose 6-phosphate (Fig. 1, ). [Note: Glucokinase has a high Km for glucose, is not subject to direct product inhibition, and has a sigmoidal reaction curve .]
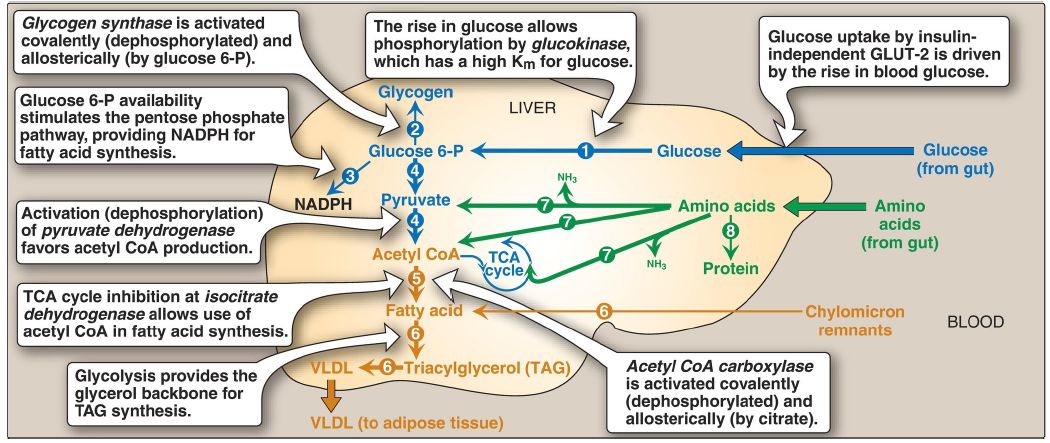
Figure 1: Major metabolic pathways in the liver in the absorptive state. [Note: The acetyl coenzyme A (CoA) is also used for cholesterol synthesis.] The numbers in circles, which appear both in the figure and in the text, indicate important pathways for carbohydrate, fat, or protein metabolism. Blue text = intermediates of carbohydrate metabolism; brown text = intermediates of lipid metabolism; green text = intermediates of protein metabolism; P = phosphate; TCA = tricarboxylic acid; VLDL = very-low-density lipoprotein; GLUT = glucose transporter; NADPH = nicotinamide adenine dinucleotide phosphate; NH3 = ammonia.
2. Increased glycogenesis: The conversion of glucose 6-phosphate to glycogen is favored by the activation of glycogen synthase, both by dephosphorylation and by increased availability of glucose 6-phosphate, its positive allosteric effector (see Fig. 1, ).
3. Increased pentose phosphate pathway activity: The increased availability of glucose 6-phosphate, combined with the active use of nicotinamide adenine dinucleotide phosphate (NADPH) in hepatic lipogenesis, stimulates the pentose phosphate pathway . This pathway typically accounts for 5%–10% of the glucose metabolized by the liver .
4. Increased glycolysis: In the liver, glycolysis is significant only during the absorptive period following a carbohydrate-rich meal. The conversion of glucose to pyruvate is stimulated by the elevated insulin/glucagon ratio that results in increased amounts of the regulated enzymes of glycolysis: glucokinase, PFK-1, and pyruvate kinase ([PK] ).
Additionally, PFK-1 is allosterically activated by fructose 2,6- bisphosphate generated by the active (dephosphorylated) kinase domain of bifunctional PFK-2. PK is dephosphorylated and active. Pyruvate dehydrogenase (PDH), which converts pyruvate to acetyl CoA, is active (dephosphorylated) because pyruvate inhibits PDH kinase (see Fig. 1, ). The acetyl CoA either is used as a substrate for fatty acid (FA) synthesis or is oxidized for energy in the tricarboxylic acid (TCA) cycle. (See Fig. 2 for the central role of glucose 6-phosphate.)

Figure 2: Central role of glucose 6-phosphate in metabolism. [Note: The presence of glucose 6-phosphatase in the liver allows the production of free glucose from the glucose 6-phosphate produced in glycogenolysis and gluconeogenesis.] NADPH = nicotinamide adenine dinucleotide phosphate; P = phosphate.
5. Decreased glucose production: While glycolysis and glycogenesis (pathways that promote glucose storage) are being stimulated in the liver in the absorptive state, gluconeogenesis and glycogenolysis (pathways that generate glucose) are being inhibited. Pyruvate carboxylase (PC), which catalyzes the first step in gluconeogenesis, is largely inactive because of low levels of acetyl CoA, its allosteric activator .
[Note: The acetyl CoA is being used for FA synthesis.] The high insulin/glucagon ratio also favors inactivation of other gluconeogenic enzymes such as fructose 1,6-bisphosphatase . Glycogenolysis is inhibited by dephosphorylation of glycogen phosphorylase and phosphorylase kinase. [Note: The increased uptake and decreased production of blood glucose in the absorptive period prevents hyperglycemia.]



|
|
|
|
التوتر والسرطان.. علماء يحذرون من "صلة خطيرة"
|
|
|
|
|
|
|
مرآة السيارة: مدى دقة عكسها للصورة الصحيحة
|
|
|
|
|
|
|
نحو شراكة وطنية متكاملة.. الأمين العام للعتبة الحسينية يبحث مع وكيل وزارة الخارجية آفاق التعاون المؤسسي
|
|
|