
























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات


Flagella—Prokaryotes
 المؤلف:
اعداد المرجع الالكتروني للمعلوماتية
المؤلف:
اعداد المرجع الالكتروني للمعلوماتية
 المصدر:
almerja.com
المصدر:
almerja.com
 الجزء والصفحة:
الجزء والصفحة:
 10-5-2016
10-5-2016
 3995
3995




+
-
20
Flagella—Prokaryotes
Flagella are the propulsive organelles of a wide variety of unicellular organisms, both prokaryotes and eukaryotes. However, prokaryotic flagella differ in almost every respect from their eukaryotic counterparts (see Flagella—eukaryotes).
1. Motility and Behavior
Prokaryotic flagella are largely external appendages that project far from the cell surface (Fig. 1, top right); for example, a typical rod-shaped bacterium is about 1 or 2 µm long, but its flagellar filaments can be 10 µm long. The motor resides at the cytoplasmic membrane and is a rotary device that transmits the rotation to the external flagellar filament. The motor is not driven by ATP hydrolysis like most mechanoenzymes, but by a proton-motive force (or in some marine and alkaliphilic bacteria, by a sodium-motive force); protons that have been actively pumped out of the cell by the electron transport chain are allowed to flow down their electrochemical potential gradient through the flagellar motor and surrender their potential energy in return for hydrodynamic work against the resistance of the aqueous medium. The filament is very thin (about 20 nm in diameter) compared to its length, with a length/diameter ratio of about 500, and has a completely regular helical or corkscrew shape. When rotated by the motor, it therefore functions as a propeller or Archimedes screw. Depending on the handedness of the helix (left-handed or right-handed) and the direction of rotation of the motor)counterclockwise or clockwise), the cell will experience a pushing or a pulling force. The flagellar motors of most bacteria are reversible, that is, they possess a switch or gear that will alternate the direction of rotation between counterclockwise and clockwise rotation. In the simplest cases, such as Pseudomonas spp., which have a single flagellum at one pole of the cell, this results in a shuttling motion. In the case of species such as Escherichia coli or Salmonella, which have several flagella originating more or less randomly around the cell body, the rotation direction that corresponds to pushing results in the individual flagellar filaments coalescing into a propulsive bundle to produce the motion called running or swimming, while the opposite direction causes the bundle to fly apart and produce a chaotic motion called tumbling. This tumbling randomly orients the cell, so that the next episode of swimming is largely uncorrelated with the previous one. The resulting trajectory is a zigzag or random walk in three-dimensional space.
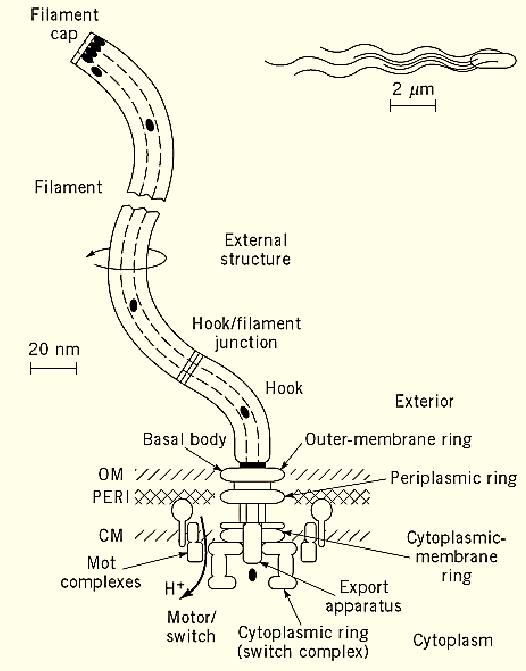
Figure 1. The bacterial flagellum, showing its four major structural elements: external structure (white), basal body) light gray), motor/switch (medium gray), and export apparatus (dark gray), along with their various substructures. The outer membrane (OM), periplasmic space (PERI), and cytoplasmic membrane (CM) of the bacterium, in which the flagellum machinery is embedded, are also shown. To drive motor rotation, protons flow through the motor down their potential gradient (toward lower H+ concentration). External proteins, such as flagellin (black ovals), are selectively passed through the export apparatus, travel down a central channel in the nascent structure, and assemble at its distal end. Most of the flagellum is tiny, so that only the filaments are evident at the scale of the bacterial cell (top right).
The above description applies when the cell is moving in an isotropic environment. However, if there are gradients of various chemicals (beneficial ones such as the amino acid aspartate or noxious ones such as phenol) and if the cell has receptors to detect these chemicals (or light, pH, temperature, etc.), it carries out an ongoing evaluation of whether its environment is improving or deteriorating. If the former is correct, it keeps going; if the latter is correct, it tumbles and then reevaluates whether its new direction is better than its old one. The net effect is a statistical drift to a better life; this behavior is called chemotaxis (or, more generally, taxis). The evaluation is in time, even though it is achieved by travel through space—a stationary bacterium could not make the measurement. Thus bacteria possess a rudimentary memory.
2. The Flagellar Gene System
Flagellar, motility, and chemotaxis genes (about 60 in all, depending on definition) are highly clustered into just four main regions of contiguous genes (Fig. 2). The organization of the genes shown in Figure 2 is for Salmonella (and is almost identical for E. coli); in other species, such as the developmental bacterium Caulobacter crescentus or the Gram-positive bacterium Bacillus subtilis, the genetic organization is somewhat different. The four gene clusters contain operons ranging in size from a single gene to more than 20 in some species. These operons fall into a hierarchy of expression by transcription, with gene products at one level playing a role in transcription of lower levels. In most cases, the controlling elements are of a more or less conventional sort, such as transcriptional activators and dedicated sigma factors for transcription initiation. However, there are at least two unconventional mechanisms. The first is a negative regulator, FlgM, which operationally seems like a repressor but is in fact an anti-sigma factor rather than a DNA-binding protein. The second is the highly unusual mechanism by which FlgM is inactivated in the last stage of the expression hierarchy; when flagellar assembly has proceeded to a fairly advanced degree, the flagellar structure itself becomes competent to export FlgM from the cell (thus inactivating it), using the same export pathway that is used for the process of flagellar export/assembly itself.

Figure 2. The four clusters of flagellar genes on the chromosome of Salmonella, along with their positions (Cs, centrism the operons (open arrows) and the genes they contain. Different gene symbols (flg, flh, and fli) are used for the four region1, Region II, which contains flagellar genes, also contains motility (mot ) genes.
3. Flagellar Structure and Function
Flagellar structure can be divided into several categories (Fig. 1):
1. External structure. This is the structure that is most evident by light or electron microscopy and includes the helical filament and a hook-shaped structure that acts as a universal joint for the filament.
2. Basal body. This structure is embedded in the cell surface (outer membrane, periplasmic space, and cytoplasmic membrane) and consists of (a) a rod whose distal end is joined to the hook, (b) an outer-membrane ring and a periplasmic ring, which together act as a bushing for the rod, and (c) a cytoplasmic-membrane ring to which the proximal end of the rod is firmly attached. The external structure and the basal body are passive components; that is, they are not part of the energy transducing machinery.
3. Motor/switch. The motor/switch can be subdivided into two categories. The first is a set of so-called Mot complexes that surround the cytoplasmic-membrane ring; these complexes, which are stationary, are involved in the generation of torque but not in the switching of rotation direction. The second is a cylindrical switch complex or cytoplasmic ring, mounted onto the cytoplasmic-membrane ring and projecting into the cytoplasm. The switch complex is involved both in the generation of torque (together with the Mot complexes) and in the switching of rotation direction. It is the rotating element of the motor and causes the cytoplasmic-membrane ring, the rod, the hook, and the filament to rotate as a single unit.
4. The export apparatus. This is the least understood structure, but available evidence suggests that it is located at the center of the cytoplasmic-membrane ring and projects out into the cytoplasm. There may also be components that shuttle between the cytoplasm and the components associated with the membrane.
The protein composition and approximate subunit stoichiometry of most of the substructures of the flagellum are known:
1. filament cap (FliD), ~10 subunits;
2. filament (FliC or flagellin), ~20,000 subunits;
3. hook/filament junction (FlgK/FlgL), ~10 subunits each.
4. hook (FlgE), ~130 subunits;
5. rod (FlgB, FlgC, FlgF, FlgG, FliE), ~5 to 25 subunits each;
6. outer-membrane ring (FlgH), ~25 subunits;
7. periplasmic ring (FlgI), ~25 subunits;
8. cytoplasmic-membrane ring (FliF), ~25 subunits;
9. cytoplasmic ring or switch complex (FliG, FliM, FliN), ~30 to 40 subunits each;
10. Mot complexes (MotA, MotB), ~12 subunits each;
11. export apparatus (composition and stoichiometry not well-characterized).
Thus there is a wide range of stoichiometries, with flagellin being by far the most abundant.
4. Flagellar Morphogenesis and the Flagellar Protein Export Process
The assembly of the flagellum proceeds for the most part in a linear fashion—that is, subunit by subunit of the first substructure, then subunit by subunit of the second substructure onto the first, and so on. The first substructures to be assembled are those associated with the cytoplasmic membrane, and the last are the ones furthest away from the cell. Thus a description (oversimplified) of the order would be: cytoplasmic-membrane ring and export apparatus → switch complex and Mot complexes → basal–body rod → periplasmic and outer-membrane rings → hook → filament.
The subunits of the basal-body rod, the hook, the filament, and some other minor but essential substructures are added to the distal end of the growing structure and get there by first passing through the export apparatus (in a process that probably requires energy and certainly requires specificity of recognition) and then traveling down a central core that exists in the structure (Fig. 1).
4.1. The Flagellar Export Pathway is a Type III Secretory Pathway
It has recently become evident that the pathway for export of flagellar proteins is but one member, albeit a rather special one, of a large family of pathways that are used by pathogenic bacteria for the secretion of virulence factors directed against the host; these pathways are called type III secretory pathways. The similarities within this family extend beyond operational ones, such as lack of signal peptide cleavage, to biochemical ones. There are at least six flagellar components (FlhA, FlhB, FliI, FliP, FliQ, FliR) that have homologues in the virulence factor secretory apparatus. FliI is an ATPase, which could be driving active transport of the protein substrates. Remarkably, it is homologous to a fundamentally important protein for bacteria (and for mitochondria and chloroplasts): the catalytic subunit of the proton-translocating F0F1-ATPase.
 0
0
 0
0
لا توجد تعليقات بعد
ما رأيك بالمقال : كن أول من يعلق على هذا المحتوى
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















