Photosynthetic O2 production
Key points: Biological solar energy capture by photoactive centres results in the generation of species with sufficiently negative reduction potentials to reduce CO2 to produce organic molecules; in higher plants and cyanobacteria, the electrons are derived from water, which is converted to O2 by a complex catalytic centre containing four Mn atoms and one Ca atom. Photosynthesis is the production of organic molecules using solar energy. It is conveniently divided into the light reactions (the processes by which electromagnetic energy is trapped) and the dark reactions (in which the energy acquired in the light reactions is used to convert CO2
and H2O into carbohydrates). We have already mentioned the most important of the dark reactions, the incorporation of CO2 into organic molecules, which is catalysed by rubisco. In this section, we describe some of the roles that metals play in the light reactions. The basic principle of photochemical energy capture, applied in a technology to produce H2 from water, was described in Chapter 10 (Box 10.3). We can view photosynthesis in an analogous way in that H2 is ‘stored’ by reaction with CO2. In biology, photons from the Sun excite pigments present in giant membrane-bound proteins known as photosystems. The most important pigment, chlorophyll, is a Mg complex that is very similar to a por phyrin (8). Most chlorophyll is located in giant proteins known as light-harvesting antennae, the name perfectly describing their function, which is to collect photons and funnel their energy to enzymes that convert it into electrochemical energy. This energy conversion uses further chlorophyll complexes that become powerful reducing agents when excited by light. Each electron released by excited chlorophyll travels rapidly down a sequence of protein-bound acceptors, including FeS clusters, and (through the agency of ferredoxin and other redox enzymes) is eventually used to reduce CO2 to carbohydrate. Immediately after releasing an electron, the chlorophyll cation, a powerful oxidant, must be rapidly reduced by using an electron from another site to avoid wasting the energy by recombination (simple reversal of electron flow). In ‘oxygenic’ photosynthesis, which occurs in green algae, cyanobacteria, and most importantly in green plants, each such ‘restoring’ electron is provided from a water molecule, resulting in production of O2. In green plants, photosynthesis occurs in special organelles known as chloroplasts. Plant chloroplasts have two photosystems, I and II, operating in series, that allow low-energy light (approximately 680–700 nm,>1 eV) to span the large potential range (>1 V) within which water is stable. The arrangement of proteins is depicted in Fig. 27.39. Some of the energy of the photosynthetic electron transfer chain is used to generate a transmembrane proton gradient which in turn drives the synthesis of ATP, as in mitochondria. Photosystem Ilies at the low-potential end, its electron donor is the blue Cu protein plastocyanin that has been reduced using the electrons generated by photosystem II; in turn, the electron donor to photosystem II is H2O. Thus green plants dispose of the oxidizing power by converting H2O into O2. This four-electron reaction is remarkable because no intermediates are released. The catalyst, called the ‘oxygen evolving centre’ (OEC) also has a special significance because its action, commencing over 2 Ga ago, has provided essentially all the O2 we have in the atmosphere. The OEC is the only enzyme active site known to produce an
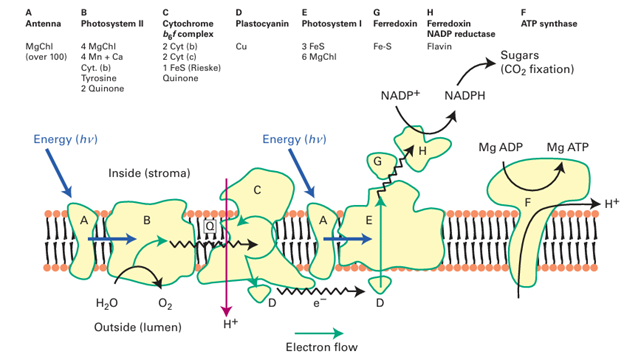
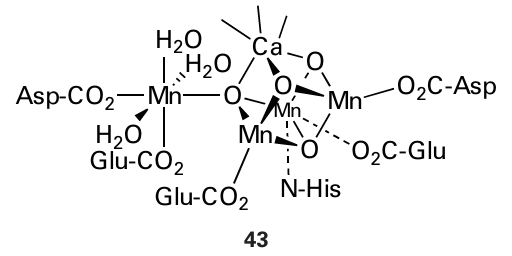
Figure 27.39 The arrangement of proteins in the photosynthetic electron-transport chain (The Mg— chlorophyll complex is represented ‘MgChl’.) A. Antenna (‘light harvesting’) complex. B. Photosystem II. C. The ‘cytochrome b6 f complex’ (this is similar to complex III in the mitochondrial ET chain). D. Plastocyanin (soluble). E. Photosystem I. F. ATPase. G. Ferredoxin (Fe S). H. Ferredoxin–NADP reductase (flavin). Blue arrows show transfer of energy. Note how the overall transfer of electrons is from Mn (high potential) to FeS (low potential): this apparently ‘uphill’ flow reflects the crucial input of energy at each photosystem. O−O bond from two H2 O molecules, and there is much interest in producing functional models of this catalyst for photochemical water splitting (Box 10.3). The OEC is a metal oxide cluster, containing four Mn atoms and one Ca atom, that is located in subunit D1 of photosystem II. Subunit D1 has long attracted interest because the cell replaces it at frequent intervals as it quickly becomes worn out by oxidative damage. X-ray diffraction data indicate that the metal atoms are arranged as a [3MnCa4O] cubane connected to a fourth ‘dangling’ Mn (43). The OEC exploits the oxidizing abilities of Mn (IV) and Mn(V), coupled with that of a nearby tyrosine residue, to oxidize H2O to O2. Successive photons received by photosystem II result in the OEC being progressively oxidized (the acceptor, an oxidized chlorophyll known as P680, has a reduction potential of approximately 1.3 V) through a series of states designated S0 to S4, as shown in Fig. 27.40. Apart from S4, which rapidly releases O2 and has not been isolated, these states are identified in kinetic studies by their characteristic spectroscopic properties, for example S2 shows a complex multi-line EPR spectrum. Note that the Mn ligands are hard O-atom donors and Mn (III) (d4), Mn (IV) (d3), and Mn(V) (d2) are hard metal ions. Based on the available structural evidence, different models have been proposed for the mechanism of O2 evolution from two H2O molecules. The overriding barrier to O2 formation lies in forming the weak peroxidic O−O bond, following which the formation of O-O is energetically easy (Section 16.1 and Resource section 3). First, as the Mn sites are progressively oxidized, coordinated H2O molecules become increasingly acidic and lose protons, progressing from H2O through OH to O2. Second, computational studies suggest that Mn(V) O is best regarded as Mn (IV)−O , in which the oxido ligand is electron deficient and has appreciable radical character. An H2O or OH ligand coordinated at one Mn subsite (or the Ca) could attack such an electron-deficient O-ligand, perhaps formed at the ‘dan gling’ Mn that is not part of the cubane. Such an attack by coordinated H2 O or OH on Mn (IV) O would result in a Mn (III)-peroxide species that is easily converted to O2 by using the reservoir of oxidizing power that has accumulated on the cluster. The presence of the Ca2 is essential, and the only metal ion that can be substituted is Sr2. A possible role for the Ca is that it provides a site that will remain permanently in the 2-oxidation state and provide a rapid and stable binding site for incoming H2O, whereas if this subsite were occupied by a fifth Mn atom, the latter would certainly become oxidized and the advantage would be lost.