![]()



النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الحيوية
مواضيع عامة في المضادات الحيوية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

حل مشكلة نهاية التضاعف End replication problem
 المؤلف:
د. محمد باقر صاحب الشهيب , أ.د. علي حمود السعدي , أ.د. حيدر كامل زيدان
المؤلف:
د. محمد باقر صاحب الشهيب , أ.د. علي حمود السعدي , أ.د. حيدر كامل زيدان
 المصدر:
مبادئ الوراثة الجزيئية
المصدر:
مبادئ الوراثة الجزيئية
 الجزء والصفحة:
ص88-90
الجزء والصفحة:
ص88-90
 2025-02-19
2025-02-19
 259
259




تواجه جزيئات الـ DNA الدائرية والخطية مواقف مختلفة عند نهاية عملية تخليق الـ DNA. ففي حالة الجزيئات الدائرية فإنها تمتلك مواقع DNA تسمى مواقع الانتهاء (ter) regions of termination في كروموسوم بكتريا القولون وتبعد 180 درجة عن منشأ التضاعف (origin of Escherichia coli chromosome (OriC بالكروموسوم الدائري. وهنالك ستة تسلسلات أخرى تتركز على كلا جانبي نقطة اللقاء وتعمل هذه التسلسلات كمنهيات terminators عندما ترتبط ببروتين الانتهاء termination protein وتأمر هذه المواقع المضاعف replisome بأن يقف ويكف عن القيام بعمله.
في حالة الجزئيات الدائرية ، فأن شوكتا التضاعف تترك منشأ التضاعف وتنتقل إلى الاتجاهات المتقابلة بعيداً عن بعضها البعض وحتماً تتقابلان في منتصف الطريق حول الدائرة. وإذا كانت أحدى الشوكتان متأخرة بالنسبة للأخرى فأنها سوف تقف عندما تصل موقع الانتهاء التابع لها وتنتظر وصول شريكتها إن بروتين الانتهاء يقوم بتثبيط إنزيم الhelicase المستخدم خلال عملية تخليق الـ DNA ولهذا يتعثر تقدم الشوكتان. وعندما تلتقي الشوكتان فهذا يعني انتهاء عملية التضاعف أما في حالة الجزيئات الخطية، ففي الكائنات حقيقية النواة كما في البشر والحيوانات ، والنباتات، والخمائر، والفطريات فكلها تمتلك كروموسومات خطية، لا تستطيع عملية التضاعف بمفردها أن تضاعف نهايات الكرموسومات الخطية بشكل كامل. وهناك طريقة خاصة تتعلق بسلوك التيلومير telomere استخدمت لضمان عدم فقدان المعلومات من نهايات الكرموسومات الخطية.
توصف عملية تضاعف الـ DNA بأنها شبه أو نصف غير مستمرة semi- discontinuous وهذا يوضح اختلاف آلية تضاعف الـ DNA الحاصلة في الشريط القائد leading strand عن تلك الحاصلة في الشريط المتلكئ lagging strand . إن الشريط القائد يتضاعف بشكل مستمر. ولكي يتضاعف الشريط المتلكئ فإن عملية بلمرة الـ DNA polymerization تبدأ من عدة بوادئ من نوع الـ RNA والتي تستطيل لتخلق قطع اوكازاكي وأخيرا تتكسر هذه البوادئ وتستبدل بتسلسلات من الـ DNA. أن إزالة أي بادئ من نوع RNA في الشريط القائد يؤدي إلى ترك فجوة والتي تملئ عادة بامتداد قطعة اوكازاكي التالية، وفي اللبائن هنالك مشكلة في تضاعف النهايات القاصية extreme ends للكروموسومات الخطية، ففي بداية السبعينات من القرن الماضي اكتشف Watson عام 1972 بأن خواص عملية تضاعف الـ DNA تمنع الخلية من نسخ نهايات الـ DNA الخطي بشكل كامل ، والتي تدعى بالتيلوميرات. وبسبب طبيعة تخليق شريط الـ lagging، فإن أنزيم بوليمريز الـ DNA لا يستطيع أن يضاعف النهاية 3' لشريط الـ DNA الخطي ذو الحلزون المزدوج بشكل كامل .عندما استنتج Watson هذه المشكلة في عام 1972 ، أوضح بأنه عندما يصل إنزيم بوليمريز الـ DNA إلى نهاية جزئية الـ DNA الخطية فسوف تكون هنالك مشكلة في إكمال التضاعف (شكل 1) عند غياب التيلومير، فأن تضاعف الـ DNA الشبه غير مستمر سوف يسبب في أنتاج جزئية خطية والتي تصبح أقصر وأقصر مع كل تضاعف. لقد بين Watson هذا التقصر بشكل غير مباشر إذ لاحظ حجب جزء من التيلومير عن فعالية إنزيم بوليمريز الـ DNA وبتلك الوسيلة يتم اعتراض القمم عن التضاعف مع كل انقسام خلوي متعاقب ، وبمعنى آخر ينقص طول التيلوميرات مع كل دورة تضاعف، ولسنوات عديدة لوحظ بأن الفقدان المتدرج للتيلوميرات ربما يعمل كآلية محفزة أساسية لبدء الشيخوخة. ولكن تنشيط إنزيم التيلومريز (وهو عبارة عن معقد من البروتينات والـ RNA، حيث يعمل المكون RNA الدور الأساسي في تحفيز إضافة التسلسلات التيلوميرية في كل دورة تضاعف بينما يقتصر دور البروتينات الأخرى على مساعدة الـ RNA في هذه العملية) في الخلايا السرطانية غالباً يعمل على تعويض ذلك الخلل بإضافة تسلسلات متلاحقة متكررة ترادفيا (TTAGGG) بعد كل عملية تضاعفية وبالتالي تحافظ الخلايا السرطانية على ديمومتها ( شكل 1 ) .
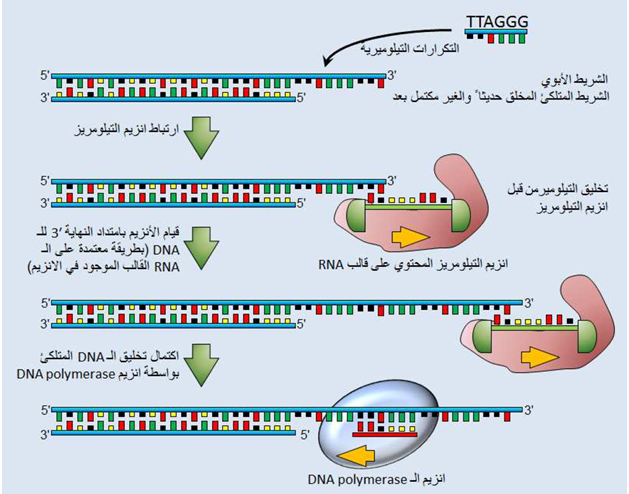
شكل (1): تضاعف التيلومير. يلخص الشكل التفاعلات الداخلة في تكوين التسلسلات الغنية بالكوانين المتكررة -G rich sequences والتي تكون نهايات الكروموسومات (التيلوميرات) لعدة كائنات حقيقية النواة. أن الشريط الغير كامل المصنع حديثاً هو الشريط المصنع في الجانب المتخلف lagging side لشوكة التضاعف. وكما هو موضح في أعلاه ،يتألف إنزيم التيلومريز من معقد RNA-protein والذي يحمل الـ RNA القالب لتخليق تسلسل الـ DNA التيلوميري الغني بالكوانين والتي هي TTAGGG في البشر. ولقد افترض في هذا التصميم أن شريط الـ lagging يتم إكماله بواسطة إنزيم DNA polymerase α والذي يحمل فعالية الـ primase في إحدى وحداته الثانوية (تصميم المؤلف)
إن هنالك عدة تجارب مهدت لاكتشاف إنزيم الـ telomerase بدأت مع ملاحظة إن فقدان التسلسلات الجينومية في كل دورة تضاعف من الممكن ان تعوض عن طريق إضافة تسلسلات طرفية، وعلاوة على ذلك تمتلك الكائنات الحية القابلية على نقل تسلسلات طرفية مختصة بالنوع إلى الـ DNA.
لقد اكتشف كل من Blackburn Elizabeth و Carol Greider في سنة 1985 بأن هنالك فعالية في مستخلصات الطفيلي Tetrahymena وتقوم هذه الفعالية بإضافة التكرارات التيلوميرية لبوادئ الـ DNA التيلوميرية قليلة النيوكليوتيدات المفردة الشريط، ووجدوا أيضا بأن هذه العملية تثبيط بمعاملة المستلخص بالإنزيم المحطم للرنا، ولهذا تسمى هذه الفعالية المعتمدة على الـ DNA بأنزيم terminal transferase أو بالتيلومريز ، حيث تحدث العملية بوساطة نسخ تسلسل الشريط القالب والذي هو جزء من المكون رنا بهذا الإنزيم. وفيما بعد وجد بأن هذا الإنزيم يتكون من المكون رنا، والفعالية الإنزيمية لهذا الإنزيم تحتاج إلى كل من المكون رنا والمكونات البروتينية. درس هذا الإنزيم بشكل مفصل باستخدام الكائن الهدبي T. thermophila لأن خلية مفردة لهذا الكائن الحي تمتلك أكثر من 40,000 تيلومير. وعموما، يمكن تلخيص آلية تفاعل إنزيم التيلومريز الرئيسية بالآتي: تمييز البادئ ، إضافة النيوكليوتيدات ، والانتقال من مكان إلى آخر (شكل 1) وبهذه الطريقة يقوم هذا الإنزيم بعد النتوء المنتهي بالنهاية 3 الغني بالكوانين والموجود بنهايات التيلوميرات.


















 جاهزية الاستعداد لشهر رمضان
جاهزية الاستعداد لشهر رمضان إستعراض موجز لحياة السيدة زينب الكبرى
إستعراض موجز لحياة السيدة زينب الكبرى أم البنين .. صانعة الوفاء وراعية الفضيلة
أم البنين .. صانعة الوفاء وراعية الفضيلة

















