![]()



النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الحيوية
مواضيع عامة في المضادات الحيوية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

كيفية عمل إنزيم التضاعف الرئيس في بكتريا القولون
 المؤلف:
د. محمد باقر صاحب الشهيب , أ.د. علي حمود السعدي , أ.د. حيدر كامل زيدان
المؤلف:
د. محمد باقر صاحب الشهيب , أ.د. علي حمود السعدي , أ.د. حيدر كامل زيدان
 المصدر:
مبادئ الوراثة الجزيئية
المصدر:
مبادئ الوراثة الجزيئية
 الجزء والصفحة:
ص73-76
الجزء والصفحة:
ص73-76
 2025-02-08
2025-02-08
 285
285




سنركز الاهتمام على إنزيم DNA polymerase III بوصفه إنزيم التضاعف الرئيسي في الخلية. إن هذا الإنزيم هو بروتين في غاية التعقيد ذو وزن جزيئي عالي 600 KDa) ) ، ويتألف من عشرة وحدات ثانوية 10 different subunits. إن ما يعرف بإنزيم البوليمريز الصميمي core polymerase يتألف من ثلاث وحدات ثانوية. تحتوي وحدة ألفا الثانوية α subunit (التي تمثل أصابع اليد fingers) على الموقع الفعال لإضافة النيوكليوتيدات بسبب امتلاكها الفعالية المبلمرة من 5' إلى 3' ووحدة ابسلون الثانوية ε subunit (التي تمثل راحة اليد palm) والتي تضطلع بإزالة النيوكليوتيدات المغلوطة mismatched nucleotides (المضافة بشكل خاطئ) بواسطة آلية معينة تعرف بآلية إصلاح الخطأ proofreading mechanism السابقة الذكر. تقوم هذه الوحدة الثانوية بهذا الدور بسبب امتلاكها الفعالية الإنزيمية المزيلة للنيوكليوتيدات والتي تعرف بـ exonuclease activity ذات الاتجاه المعاكس لعملية البلمرة والذي هو من 3' إلى 5'. أما الوحدة الثانوية الثالثة والتي تعرف بثيتا subunit θ( والتي تمثل الإبهام thumb) فهي على أية حال غير معروفة الوظيفة (شكل 1 ) .

شكل (1) تركيب انزيم البوليميريز الصميمي core DNA polymerase في بكتيريا القولون حسب معطيات تقنية X-rays crystallography وببساطة يشبه انزيم DNA polymerase اليد اليمنى نصف المفتوحة ، حيث يلاحظ ان كل من راحة اليد palm ، والاصابع fingers ، والابهام thumb يقبض على الDNA بالإضافة الى ذلك ، يبرز موقعين منفصلين مكانياً عن بعضهما وهما موقع البلمرة وموقع التدقيق في الانزيم ( تصميم المؤلف ) .
يتقدم إنزيم DNA polymerase III كما نوقش سابقا - بالاتجاه من 5' إلى 3' وذلك بسبب امتلاكه لوحدة α ذات الفعالية المبلمرة من 5' إلى 3' ولا يمكن له من أن يتقدم بالاتجاه المعاكس بسبب عدم امتلاكه لوحدة ثانوية أخرى ذات فعالية بلمرة من 3' إلى 5'. ولكن، يحدث في بعض الأحيان أن تندمج نيوكليوتيدة مغلوطة في سلسلة الـ DNA النامية، وفي هذه الحالة، يتوقف إنزيم DNA polymerase III عن العمل، ثم ينقل النهاية 3' للسلسلة النامية من وحدة α المبلمرة (أصابع اليد)، إلى وحدة ε الهاضمة ذات الاتجاه المعاكس. وهكذا تزال النيوكليوتيدة المغلوطة بالاتجاه من 3' إلى 5'. وحالما تنتهي عملية إزالة النيوكليوتيدة المغلوطة بآلية تصحيح الخطأ المعاكسة لاتجاه عملية البلمرة ترجع النهاية '3 لسلسلة الـ DNA النامية إلى موقع البلمرة في وحدة α ، وتستأنف عملية البلمرة بالاتجاه من 5' إلى 3'( شكل 1) .
أما الدور الرئيسي للوحدات الثانوية المتبقية (6 وحدات ثانوية) هو تحويل إنزيم البوليمريز الصميمي من إنزيم تفريقي distributive (غير تقدمي)، أي الإنزيم الذي يسقط من الشريط القالب بعد تخليقه من عشر الى خمسين نيوكليوتيدة، إلى إنزيم تقدمي processive والذي يمكن له تخليق امتدادات طويلة من الـ DNA تصل إلى أكثر من 500 ألف نيوكليوتيدة بدون أن يسقط من الشريط القالب. إن الفعالية الأخيرة تكون ضرورية للتخليق الكفء لكلا شريطي الـ leading مع شريط الـ lagging.
إن أساس الطبيعة التقدمية لإنزيم DNA polymerase III هو قابلية وحدة بيتا الثانوية β subunit على تكوين شكل يشبه الكعكة أو الحلقة حول الحلزون المزدوج وذلك لربط وحمل الإنزيم الصميمي (شكل2) وهنا تبرز أحد الاجابات الرئيسية عن التساؤل عن سبب تدخل انزيم معقد كأنزيم DNA polymerase III في عملية التضاعف على الرغم من امتلاك انزيم DNA polymerase I لنفس الفعالية المبلمرة التي يمتلكها الانزيم III ،بسبب عدم قدرة الانزيم I على اطالة سلسلة الـ DNA لمسافات طويلة كما هو عليه الحال في الانزيم III وهذا هو أحد الأسباب الرئيسية في تعقيد الانزيم III مقارنة بالأنزيم I ،حيث احتاج الانزيم III الى عدة وحدات ثانوية لتقوم بهذا الدور.
تعد جزيئات DNA polymerase core لوحدها قادرة على تخليق امتداد قصير فقط من النيوكليوتيدات قبل تتهاوى عن الشريط القالب. ولكن هذا لن يحصل للإنزيم بسبب وجود تركيب يعرف بالكلاب clamp والذي يحافظ على إنزيم الـ polymerase مرتبطا بقوة على شريط الـ DNA وذلك من خلال ارتباط هذا الكلاب بالـ DNA من جهة وبالإنزيم الصميمي من جهة أخرى.
إن وجود الكلاب المنزلق sliding clamp جعل لإنزيم DNA polymerase خاصيتين متناقضتين ، فعلى الرغم من اتصاله المحكم بالـ DNA وعلى امتدادات طويلة، إلا انه رخو بشكل كاف لكي يتحرك بشكل انزلاقي متسلسل من نيوكليوتيدة إلى أخرى. يرجع سبب وجود هذه الخواص المتناقضة إلى شكل وحدة β الثانوية التي تشبه الكعكة المحيطة بالـ DNA ، أو التي تشبه الحلقة التي تنزلق على الحبل (شكل 2 ).

شكل (2) وحدة بيتا الثانوية المزدوجة β-subunit dimer تقيد إنزيم E. coli DNA polymerase III الصميمي بالـ DNA، وبهذه الطريقة، تزيد من صفة التقدمية processivity لهذا الإنزيم لأكثر من ألف مرة (تصميم المؤلف).
كيف يمكن لهذا الكلاب منع إنزيم الـ polymerase من الانفصال وبنفس الوقت بدون عرقلة حركة إنزيم الـ polymerase السريعة على طول جزيئة الـ DNA؟ توصلت الدراسات الجارية على هذا الكلاب إلى أنه يكون حلقة كبيرة حول حلزون الـ DNA المزدوج. ولوحظ إن جانب واحد من الحلقة يرتبط بالجزء الخلفي من إنزيم DNA polymerase، بينما تنزلق الحلقة كاملة وبشكل حر على طول الـ DNA كلما تحرك الإنزيم. لا يتصل الكلاب من تلقاء نفسه بإنزيم الـ DNA polymerase ، إذ لا بد من وجود محمّل الكلاب clamp loader أو ما يعرف بمعقد كاما γ complex، والذي يشبه حمالة البنطلون ويتألف من خمسة وحدات ثانوية، والذي وبمساعدة التحلل المائي للـ ATP يقوم بربط الكلاب بإنزيم الـ DNA polymerase (شكل 3). وبهذه الطريقة، تزداد صفة تقدمية الإنزيم .
يتوسط معقد كاما γ complex وظيفتين مهمتين (1) تحميل load كلاب وحدة بيتا الثانوية β-subunit clamp بالإنزيم الصميمي عند بداية تخليق شريط الـ DNA، في تفاعل يحتاج إلى طاقة الـ ATP (2) تفريغ تحميل unload كلاب وحدة بيتا الثانوية بعد اكتمال تخليق شريط الـ DNA (شكل 3)
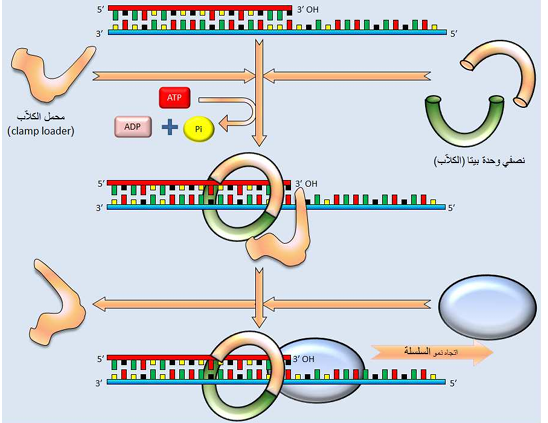
شكل (3) مخطط يوضح كيفية تراكب الكلاب لكي تبقي من انزيم الـ DNA polymerase متحركاً على الـ DNA وذلك في التفاعل المبسط المبين في أعلاه . إن محمل الكلاب clamp loader ينفصل في حال تراكب الكلاب على انزيم الـ polymerase (تصميم المؤلف ).


















 جاهزية الاستعداد لشهر رمضان
جاهزية الاستعداد لشهر رمضان إستعراض موجز لحياة السيدة زينب الكبرى
إستعراض موجز لحياة السيدة زينب الكبرى أم البنين .. صانعة الوفاء وراعية الفضيلة
أم البنين .. صانعة الوفاء وراعية الفضيلة

















