























الفاكهة والاشجار المثمرة
نخيل التمر

النخيل والتمور
آفات وامراض النخيل وطرق مكافحتها
التفاح
الرمان
التين
اشجار القشطة
الافو كادو او الزبدية
البشمله او الاكي دنيا
التوت
التين الشوكي
الجوز
الزيتون
السفرجل
العنب او الكرمة
الفستق
الكاكي او الخرما او الخرمالو
الكمثري(الاجاص)
المانجو
الموز
النبق او السدر
فاكة البابايا او الباباظ
الكيوي
الحمضيات
آفات وامراض الحمضيات
مقالات منوعة عن الحمضيات
الاشجار ذات النواة الحجرية
الاجاص او البرقوق
الخوخ او الدراق
الكرز
المشمش
مواضيع عامة
اللوز
الفراولة او الشليك
الجوافة
الخروب(الخرنوب)
الاناناس
مواضيع متنوعة عن اشجار الفاكهة
التمر هندي
الكستناء
شجرة البيكان ( البيقان )
البندق
المحاصيل
المحاصيل البقولية
الباقلاء (الفول)
الحمص
الترمس
العدس
الماش
اللوبياء
الفاصولياء
مواضيع متنوعة عن البقوليات
فاصوليا الليما والسيفا
محاصيل الاعلاف و المراعي
محاصيل الالياف
القطن
الكتان
القنب
الجوت و الجلجل
محصول الرامي
محصول السيسال
مواضيع متنوعة عن محاصيل الألياف
محاصيل زيتية
السمسم
فستق الحقل
فول الصويا
عباد الشمس (دوار الشمس)
العصفر (القرطم)
السلجم ( اللفت الزيتي )
مواضيع متنوعة عن المحاصيل الزيتية
الخروع
محاصيل الحبوب
الذرة
محصول الرز
محصول القمح
محصول الشعير
الشيلم
الشوفان (الهرطمان)
الدخن
محاصيل الخضر
الباذنجان
الطماطم
البطاطس(البطاطا)
محصول الفلفل
محصول الخس
البصل
الثوم
القرعيات
الخيار
الرقي (البطيخ الاحمر)
البطيخ
آفات وامراض القرعيات
مواضيع متنوعة عن القرعيات
البازلاء اوالبسلة
مواضيع متنوعة عن الخضر
الملفوف ( اللهانة او الكرنب )
القرنبيط او القرنابيط
اللفت ( الشلغم )
الفجل
السبانخ
الخرشوف ( الارضي شوكي )
الكرفس
القلقاس
الجزر
البطاطا الحلوه
القرع
الباميه
البروكلي او القرنابيط الأخضر
البنجر او الشمندر او الشوندر
عيش الغراب او المشروم او الأفطر
المحاصيل المنبهة و المحاصيل المخدرة
مواضيع متنوعة عن المحاصيل المنبهة
التبغ
التنباك
الشاي
البن ( القهوة )
المحاصيل السكرية
قصب السكر
بنجر السكر
مواضيع متنوعة عن المحاصيل
نباتات الزينة والنباتات الطبية والعطرية
نباتات الزينة
النباتات الطبية والعطرية
الحشرات النافعة
النحل
نحل العسل
عسل النحل ومنتجات النحل الاخرى
آفات وامراض النحل
دودة القز(الحرير)
آفات وامراض دودة الحرير
تربية ديدان الحرير وانتاج الحرير الطبيعي
تقنيات زراعية
الاسمدة
الزراعة العضوية
الزراعة النسيجية
الزراعة بدون تربة
الزراعة المحمية
المبيدات الزراعية
انظمة الري الحديثة
التصنيع الزراعي
تصنيع الاعلاف
صناعات غذائية
حفظ الاغذية
الانتاج الحيواني
الطيور الداجنة
الدواجن
دجاج البيض
دجاج اللحم
امراض الدواجن
الاسماك
الاسماك
الامراض التي تصيب الاسماك
الابقار والجاموس
الابقار
الجاموس
امراض الابقار والجاموس
الاغنام
الاغنام والماعز
الامراض التي تصيب الاغنام والماعز
آفات وامراض النبات وطرق مكافحتها
الحشرات
الحشرات الطبية و البيطرية
طرق ووسائل مكافحة الحشرات
الصفات الخارجية والتركيب التشريحي للحشرات
مواضيع متنوعة عن الحشرات
انواع واجناس الحشرات الضارة بالنبات
المراتب التصنيفية للحشرات
امراض النبات ومسبباتها
الفطريات والامراض التي تسببها للنبات
البكتريا والامراض التي تسببها للنبات
الفايروسات والامراض التي تسببها للنبات
الاكاروسات (الحلم)
الديدان الثعبانية (النيماتودا)
امراض النبات غير الطفيلية (الفسيولوجية) وامراض النبات الناتجة عن بعض العناصر
مواضيع متنوعة عن امراض النبات ومسبباتها
الحشائش والنباتات الضارة
الحشائش والنباتات المتطفلة
طرق ووسائل مكافحة الحشائش والنباتات المتطفلة
آفات المواد المخزونة
مواضيع متنوعة عن آفات النبات
المكائن والالات الزراعية
مواضيع متنوعة عن الزراعة

هيكلية فايروسات النبات
 المؤلف:
نبيل عزيز قاسم
المؤلف:
نبيل عزيز قاسم
 المصدر:
فايروسات النبات (2011م)
المصدر:
فايروسات النبات (2011م)
 الجزء والصفحة:
ص 66-88
الجزء والصفحة:
ص 66-88
 2-8-2017
2-8-2017
 6545
6545




+
-
20
هيكلية فايروسات النبات
هيكلية فايروسات النبات Architecture Of plant Viruses هو النظام الذي تترتب به الوحدات البنائية البروتينية لتكوين الكابسيد والكيفية التي يرتبط بها الحامض النووي بتلك الوحدات ونوعية تناظر جسيمات الأنواع الفايروسية، ولغرض فهم هيكلية فايروسات النبات بتنوع أشكالها فانه يجب معرفة طبيعة الكابسيد الفايروس بأنواعه المختلفة وحسب شكل الفايروس.
1. الكابسيد (الغطاء البروتيني)
الكابسيد Capsid هو الغطاء البروتيني للفايروسات Coat or Protein Shell وهو تركيب عالي التنظيم يتكون من تجمع هندسي للوحدات البنائية البروتينية يحيط ويرتبط بالحامض النووي وذلك في كل أنواع فايروسات النبات وبذلك يتكون من ارتباطهما الفيزيائي "النيوكليوكابسيد" Nucleocapsid وهو المكون المعقد المتكون من ارتباط الكابسيد بالحامض النووي والذي يمثل كيميائيا "النيوكليوبروتين" Nucleoprotein وهذان المصطلحان يعبران كيماويا عن الفايروسات النباتية البسيطة، أما في الفايروسات المغلفة فيطلق على النيوكليوكاسيد مصطلح "اللب" Core والذي يغلف بغلاف ليبوبروتيني إضافي. إن أول من استعمل مصطلح الكابسيد هو لوف" (Lowff) في نهاية خمسينات القرن العشرين، وعرفه Casper وآخرون (1962) بأنه الغطاء البروتيني المتناظر الذي يضم الحامض النووي الفايروسي مكونا النيوكليوكابسيد، ويتكون من وحدات أساسية هي الوحدات البنائية التركيبية. يتكون كابسيد الفايروسات البسيطة والتي تشكل النسبة الأكبر من فايروسات النبات من وحدات بنائية متماثلة وذلك بسبب محدودية أعداد الجينات الفايروسية مما لا يسمح بتنويع الوحدات البنائية حيث أن تنويع وتعقيد هذه الوحدات يتطلب المزيد من المعلومات الوراثية (الجينات) كذلك فان هذا التماثل منح الفايروسات ميزة التجميع الذاتي للوحدات البنائية Self assembly عند بناء الكابسيد للوحدات البنائية أي ترتيب تلك الوحدات بطريقة لا تسمح بالخطأ عند البناء حيث ترفض أية وحدة بنائية لا تنسجم مع الترتيب المطلوب وانعكس ذلك على محدودية تصاميم الأشكال الفايروسية التي انحصرت في الأشكال الخمسة المذكورة، إن هذه المحدودية في الأشكال الفايروسية قد تبدو سلبية تعانيها الفايروسات ولكنها في الواقع ميزة لها في اتجاهين (1) الاقتصاد في مكونات بناء الغطاء البروتيني أي في الوحدات البنائية (2) كفاءة البناء حيث أن الشكل البلوري هو أكفأ بناء هندسي من ناحية الاستغلال الأمثل للأسطح نسبة إلى الحجوم.
تعود محدودية الأشكال الفايروسية إلى (1) أن التركيب الثالثي للوحدات البنائية مقيد بتتابع الأحماض الامينية فيها، واقترح الباحثون نموذجا لكيفية تعاقب تلك الأحماض في الوحدة البنائية أطلق عليه "الصندوق الكاربوكسيسلي Carboxy Cage والذي تترتب ست مجاميع كاربوكسيسلية فيه (2) تأثير التركيب الرابعي للبروتين الناتج من التجميع الذاتي للوحدات البنائية عند بناء الكابسيد.
يتكون الكابسيد في الفايروسات من المكونات الأساسية التالية:
1. الوحدات البنائية التركيبية الأساسية
هي الوحدات الأساسية للكابسيد والتي يصطلح عليها Structural units أو تسمى "الوحدة الشكلية" Morphological units وهي عبارة عن سلسلة ببتيد أحادية الالتفاف Single folded peptide أي تمثل التركيب الثالثي للبروتين، وإذا ما تكونت من نوع واحد من الببتيد عندها تسمى "الوحدة الكيميائية" Chemical Unit أو "الوحدة التحتية البروتينية" Protein Subunit وهي تمثل فصوص الكابسيد في الهياكل الحلزونية والمكعبية، (الشكل 1)، يتكون كابسيد بعض أنواع الفايروسات من وحدات بنائية متماثلة كما في الفايروسات البسيطة لذا يعد الكابسيد في مثل هذه الحالة "مكوثر متجانس" Homopolymer أو Multiply redundants وذلك لتكرار نسخ متماثلة من الوحدات البنائية فيه، أما في الفايروسات الأكثر تعقيدا فيتكون الكابسيد من وحدات بنائية غير متماثلة أي مكونة من أكثر من نوع من البروتين ليكون مكوثر غير متجانس" Heteropolymer.
2. الكابسومير
الكابسومير Capsomer والذي يسمى أيضا الوحدة الشكلية Morphological Unit هو الوجه المثلثي للفايروسات الأيزومترية البلورية ويتكون من تجمع عنقودي منتظم بشكل مثلث متساوي الأضلاع لعدد من الوحدات البنائية التركيبية المتماثلة أو غير المتماثلة، (الشكلين 1 و 2)، ولا يوجد الكابسومير في الفايروسات الأنبوبية حلزونية التناظر.

الشكل (1): هيكلية الفايروسات العصوية المرنة والصلدة والايزومترية وترتيب الوحدات البروتينية الحامية للحامض النووي (أ) مخطط يبين ارتباط الحامض النووي مع الوحدات البنائية البروتينية (الفصوص) في الفايروسات العصوية المرنة والصلدة وان خيط الحامض الحلزوني الواضح أعلى الشكل هو للتوضيح أما في الواقع فانه منطمر تماما في الوحدة البنائية البروتينية الأخيرة ولا يظهر منه شيء، فيما يشير محور الجسيمة إلى محور التناظر، لا يوجد كابسومير في هذا الجسم بل إن الفصل هو الذي يمثله (ب) مخطط يبين ترتيب الوحدات البنائية البروتينية مكونة كابسوميرات خماسية وسداسية لتكوين البلورة النموذجية الفايروسية عشرينية الوجه Icosahedron ذات رقم التثليث = 3، ولا يظهر الحامض النووي لأنه معبأ داخل البلورة (ج) بلورة فايروس موزائيك الجت (AMV) المكونة من 30 كابسومير من نوع "المثنى Dimer.
الشكل مقتبس من Hull ( (2002.

الشكل (2): مخطط عام لهيكلية الفايروسات النباتية (1) فايروس عصوي مرن (2) فايروس عصوي صلد (3) قطعة توضح ارتباط الحامض النووي بالوحدات البروتينية في النوعين السابقين (4) منظر فوقي للعصية الصلدة أو المرنة يبين كيفية ارتباط الحامض النووي بالوحدات البروتينية (5) مقطع في فايروس باسيلي يظهر ترتيب خيط الحامض النووي داخل العصية (6) مقطع عرضي للعصية الباسيلية حيث تمثل الدائرة الخارجية الغطاء البروتيني والدائرة الداخلية الحامض النووي (7) ترتيب الوحدات البروتينية لكابسيد فايروس آيزومتري (8) مخطط هندسي لكابسيد فايروس آيزومتري يبين الاوجه المثلثيه الممثلة للكابسوميرات (9) ترتيب الوحدات البروتينية لكابسيد الجسيمتين الثنائيتين Geminate المكونة لفريون فايروسات Geminiviruses.
الشكل مقتبس من Agrios (.(2005
2. أنواع التناظر في فايروسات النبات
يوجد نوعين رئيسيين من التناظر لفايروسات النبات وهما:
1. التناظر الحلزوني
التناظر الحلزوني Helical Symmetry هو التناظر الثنائي الذي يميز الفايروسات الأنبوبية بنوعيها الصلد والمرن وكذلك الفايروسات الباسيلية وشبيهة الاطلاقة. والتناظر الثنائي لأي جسم يعني انتصاف ذلك الجسم إلى نصفين متماثلين تماما عند مرور خط وهمي في محوره الطولي، (الشكل 1 أ).
2. التناظر المكعبي
التناظر المكعبي Symmetry Cubic أو التناظر البلوري وهو الذي تمتاز به كل أنواع الفايروسات البلورية أو الكروية أو الايزومترية أو العشرينية الوجه وهو تناظر ثلاثي حيث تمتلك البلورة الفايروسية ثلاثة محاور تدويرية Rotational Symmetry وذلك حسب مسقط النظر عند تدوير الجسيمة البلورية وهي (1) التناظر الثنائي 2-fold Symmetry وهو الذي ينصف البلورة إلى نصفين متماثلين وذلك عند مرور خط وهمي من منتصف الحافة Edge التي تفصل كل مثلثين متجاورين لاثنين من المخمسات Pentagons المكونة للبلورة حيث أن كل مخمس مكون من خمسة مثلثات متساوية الأضلاع تلتقي في نقطة مدببة هي "السمت" Vertix وبذلك يمتلك الجسم البلوري الفايروسي 15 محورا ثنائي التناظر وعند النظر على كل منها عموديا سترى البلورة بجزئيها المتماثلين (2) التناظر الثلاثي 3-fold Symmetry هو التناظر الذي يقسم البلورة الفايروسية إلى ثلاثة أجزاء متماثلة عند مرور خط وهمي في مركز كل وجه مثلثي حيث تضم البلورة الفايروسية العشرينية الوجه عشرة محاور ثلاثية 3-fold axes عليه عند النظر عموديا على مركز كل وجه مثلثي ستظهر البلورة وكأنها مقسمة إلى ثلاثة أجزاء متماثلة (3) التناظر الخماسي 5-fold Symmetry هو التناظر الذي يقسم البلورة الفايروسية إلى خمسة أقسام متماثلة وذلك عند مرور خط وهمي من محور يمر من قمة السمت حيث تضم البلورة ستة محاور خماسية، (الشكل 3) توجد الفايروسات البلورية بأشكال مختلفة تتباين في درجة تعقيد بلوراتها وكما مبين في الشكلين (2 و 4) الذي يظهر مخططات للبلورات الفايروسية العشرينية المكونة من كابسوميرات ثنائية وخماسية وسداسية.
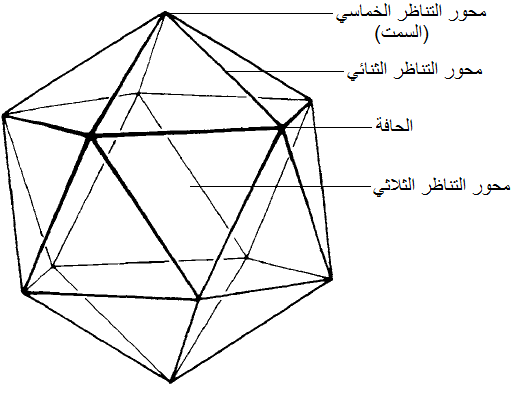

(أ) (ب)
الشكل (3): انواع التناظر في الفايروسات البلورية (أ) الشكل الهندسي لبلورة نموذجية لفايروس عشريني الاوجه Icosahedron وفيها 12 سمت و 20 وجه مثلثي متماثل يبين محاور التناظر الثلاثة وهي: التناظر الثنائي، والتناظر الثلاثي، والتناظر الخماسي (ب) مخطط توضيحي لمحاور التناظر الثلاثة.
الشكل مقتبس من Carter و Saunders (2007) و Hull (2002).


الشكل (4): أنواع من الفايروسات الأيزومترية (أ) الجسيمات البلورية لفايروس التقزم الاصفر للشعير(BYDV) (ب) فايروس تبرقش البيلادونا (BeMV) حيث يظهر الشكل الجسيمات الكاملة وعدد من الجسيمات الفارغة من الحامض النووي ذات المركز الغامق بسبب دخول الصبغة فيها، والصورة المكبرة إلى اليمين لجسيمة مكبرة تظهر فيها الكابسوميرات (ج) الجسيمات الكروية لفايروس التقزم الشجيري للعليق (RBDV) (د) الجسيمات البلورية لفايروس التبرقش الاصفر للذرة (MCMV) (ه) شكل مجسم للتركيب السطحي لفايروس موزائيك البروم يظهر الكابسوميرات الخماسية والسداسية (و) شكل مجسم لكابسيد فايروس التبرقش الاصفر للرز (RYMV) يظهر تكونه من أكثر من نوع من البروتينات وترتيب شبه مكافئ.
الشكل مقتبس من Mahy و Van Regenmortel (2008).
3. كيفية بناء الفايروسات العصوية
يعد فايروس موزائيك التبغ (TMV) أفضل مثالا لتوضيح هيكلية الفايروسات العصوية وكيفية بناؤها فجسيماته عصية حلزونية صلدة بطول وقطر 18300 X نانومتر على التوالي ويشكل البروتين 95% من كتلتها والمتبقي هو الحامض النووي، يتكون الكابسيد من 2130 وحدة بروتينية مرتبة حلزونيا Helical array بترتيب محكم التنظيم ومماثل للترتيب الحلزوني للحامض النووي بحيث ترتبط ثلاث نيوكليوتايدات مع كل تحت وحدة بروتينية من وحدات الكابسيد، وتبلغ قيمة "خطوة الحلزون" Pitch of hellx 2,3نانومتر، وخطوة الحلزون هي عدد النيوكليوتايدات في الدورة الحلزونية الواحدة والتي يصبح فيها موقع نيوكليوتايدة معينة تماما فوق مسقط النيوكليوتايدة الأولى التي بدأت بها الدورة وتقاس بالمسافة التي تشغلها دورة كاملة من الحلزون في جينوم الفايروسات العصوية وهي صفة تشخيصية لكل فايروس عصوي مرن أو صلد. تتكون دورة حلزون هذا الفايروس من 49 نيوكليوتايدة وهذا يعني أنه لكي يصبح مسقط النيوكليوتايدة الأخيرة تماما فوق مسقط النيوكليوتايدة الأولى التي بدأ بها الالتفاف فان هذا يتطلب ثلاث دورات حلزونية لكابسيد هذا الفايروس أي 49 نيوكليوتايدة و 16.3 تحت وحدة بروتينية لأن كل واحدة منها ترتبط بثلاث نيوكليوتايدات، (تحت الوحدات البروتينية لا تتجزأ ولكن المسافة التي تشغلها 49 نيوكليوتايدة تساوي مسافة 16,3 تحت وحدة بروتينية) وهكذا يتكرر هذا النظام كل ثلاث دورات لغاية نهاية العصية، (الشكل 5) تبتعد مجاميع الفوسفات بحدود 4 نانومتر عن محور العصية ويكون اتجاه دوران الحلزون هو الاتجاه اليميني Right-handed أي من اليسار إلى اليمين، وتكون إحدى نهايتي العصية مقعرة Concave ترتبط بها النهاية 5 للحامض النووي أما النهاية الأخرى فهي محدبة Convex وترتبط بها النهاية 3، ويبلغ قطر القناة المركزية المجوفة للعصبية 4 نانومتر .
تنتظم سلسلة الحامض النووي بأحكام في الحلزون متداخلة مع كل الوحدات البروتينية وتبقى منطمرة في الوحدتين النهائيتين الأولى والأخيرة من دون أن تظهرا منهما. تتكون الوحدة البروتينية لهذا الفايروس من 158 حاض أميني وتأخذ النسبة الأكبر من هذا البروتين شكل التركيب الثاني Secondary Structure حيث تكون ما يقرب من 50% من الأحماض الامينية الداخلة في بناء الوحدة البروتينية أربعة تراكيب حلزونية من النوع "الفا" α– helix، و 10% من النوع "بيتا " ß – helix فيما تشكل بقية الأحماض الامينية الدورات العكسية Reverse turns، (الشكل 6) وتكون النهايتين الكاربوكسيلية والامينية للبروتين باتجاه السطح الخارجي للجسيمة.
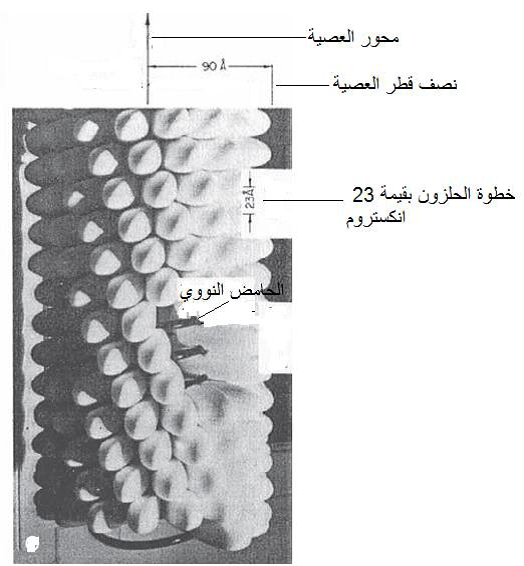
الشكل (5): مجسم توضيحي يبين هيكلية جسيمة (عصية) فايروس موزائيك التبغ (TMV) وخطوة الحلزون ونصف قطر الجسيمة بقيمة 90 انكستروم.
الشكل مقتبس من Hull (2009).
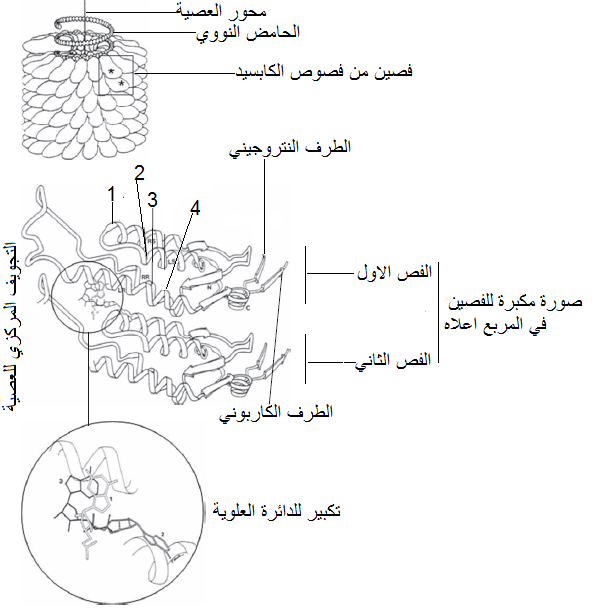
الشكل (6): تركيب كابسيد جسيمة فايروس موزائيك التبغ (TMV) وكيفية ارتباط الحامض النووي مع الوحدات البروتينية للكابسيد، يوضح المربع في الشكل العلوي إثنين من فصوص الكابسيد والمكبر في الشكل الوسطي حيث يظهر أن كل فصل يتكون من اربعة سلاسل حلزونية من نوع "الكاربون الفا" α – Carbon helix والتي تمثل التركيب الثاني للبروتين حيث يشير الرقم 1 إلى السلسلة يمينية الاستدارة Right-slewed والرقم 2 إلى السلسلة يسارية الاستدارة Left-slewed والرقم 3 إلى السلسلة اليمينية القطرية – Right radial والرقم 4 إلى السلسلة اليسارية القطرية Left- radial وتظهر العروة الطويلة Long loop اقصى اليسار والتي تربط السلسلتين القطريتين اليمينية واليسارية معا. يبين الشكل الدائري السفلي تكبيرا لمنطقة تداخل الحامض النووي مع الوحدات البروتينية حيث
ترتبط ثلاث نيوكليوتايدات مع كل تحت وحدة بروتينية من وحدات الكابسيد.
الشكل مقتبس من (2009) Hull.
1. مواقع ربط الحامض النووي الرايبي ببروتين الغطاء
يرتبط الحامض النووي في موقعين ارتباطيين مع كل وحدتين بروتينيتين متعاقبتين
وهذان الموقعان يوجدان على قمة الوحدة البروتينية الأولى وأسفل الثانية حيث ترتبط كل ثلاث قواعد نتروجينية مع وحدة بروتينية واحدة بعد أن تكون هذه القواعد المخالب" Claw الذي يمسك بإحكام بالحلزون القطري الأيسر للوحدة البروتينية العليا إذ يمتلك هذا الجزء من الحلزون العدد الأكبر من الأحماض الامينية الاليفاتية بين الموقعين 117 و128 من تسلسل الأحماض الامينية في كل وحدة بروتينية. ترتبط مجموعتي الفوسفات أيونيا مع اثنين من جزيئات الحامض الاميني "الارجنين" Arginine في الموقعين 90 و 92، فيما تكون مجموعة الفوسفات الثالثة أصرة هيدروجينية مع الحامض الاميني "الثريونين" Thrionine، (الشكل 7) ويمكن فهم طبيعة الأواصر الالكتروستاتيكية بين البروتين والحامض النووي من خلال حالة التكامل Complementarity الحاصلة بين الأسطح الالكتروستاتيكية للبروتين والحامض النووي الرايبي حيث أنها ليست مجرد أواصر أيونية بين الحامض الأميني "الارجنين" والفوسفات، ولوحظ أيضا وجود جزيئات ماء منتشرة بين الأسطح الداخلية والخارجية للوحدات البروتينية والتي تلعب دورا في تماسك الوحدات البروتينية ببعضها، ويلعب أيون الكالسيوم دورا مهما في تضخيم جسم الفريون وتجميع الوحدات البروتينية.
فشل كابسيد فايروس موزائيك التبغ تماما بالارتباط مع الدنا الفايروسي في تجارب أجريت خارج النسيج الحي in vitro ويعني ذلك انه متخصص للارتباط فقط مع الرنا الفايروسي، وتلعب مجاميع هيدروكسيل الرايبوز الحرة الدور الأهم في هذه التخصصية الارتباطية لأنها لا تسمح بالارتباط مع الثايمين. إن عملية الربط التي شرحت أعلاه هي بداية عملية تجميع الحامض النووي الفايروسي مع الكابسيد لتكوين الجسيمة الفايروسية الكاملة وكما موضح في الفقرة التالية.

الشكل (7): مخطط يبين موقع ارتباط الحامض النووي (الرنا) لفايروس موزائيك التبغ (TMV) مع الوحدات البروتينية لقرصين من الكابسيد، تشير الارقام 1 – 4 إلى ذات السلاسل الحلزونية من نوع "الكاربون ألفا" – Carbon helix α المبينة في الشكل (6) ويظهر موقع ارتباط الرنا بالأصرة الرابطة بين القرصين، وتشير الارقام الموضحة على السلاسل إلى تسلسل الأحماض الأمينية الداخلة في تركيبها. الشكل مقتبس من Hull ( (2002
2. الية تجميع الحامض النووي مع الكابسيد
درست الية التجميع فايروس موزائيك التبغ ممثلا للفايروسات العصوية والتي تتم بثمانية مراحل يبينها الشكل (8)، وتبين هذه الألية أن الكابسيد لا يرتبط بالحامض النووي بطريقة عشوائية بل بطريقة عالية التنظيم وان القوى المسئولة عن التجميع Reassembly هي الأواصر الكاره للماء والأواصر الالكتروستاتيكية. يبتدئ الارتباط (التجميع) بتكوين "القرص المزدوج Double helix البروتيني وهو مكوثر ذو معامل ترسيب S20 ويكون هذان القرصان حلقتين من 17 وحدة بروتينية وترتبط الحلقة الأولى مع التي فوقها بثلاث مناطق تماس عمودية بين الأحماض الامينية ويكون الارتباط في الجزء الخارجي الأيمن من القرص أي باتجاه خارج الجسيمة فيما تتفتح الأحماض الامينية إلى الداخل أي باتجاه التجويف المركزي للعصية وبشكل مرن لتشكل زوجا من الفكوك بشكل سلاسل ملتوية Folded chains تمثل الأجزاء الداخلية المرنة من القرص والمبينة بالخطوط المنقطة في الشكل (7) وهي التي ترتبط مع الحامض النووي بالطريقة المبينة في الفقرة السابقة.
يتكون القرص المزدوج بشكل متراكب بارتباط أطراف القرصين بما يشبه الفكة القافلة" Lock washer والتي تمثل بداية عملية التجميع بتأثير الأس الهيدروجيني والقوة الأيونية ودرجة حرارة خلية العائل حيث تتجمع الوحدات البروتينية لتشكل القرصين وذلك عندما يكون الأس الهيدروجيني لخلية العائل 6,5-7، (الشكل 9) وبعد تكوين القرص المزدوج يتم تجميعه مع الحامض النووي ويبدأ التجميع بين النيوكليوتايدتين 900-1300 ومن النهاية 3 حيث يلتف الحامض النووي مكونا عروة تبدأ بالالتحام مع وحدات البروتين بدخولها في التجويف المركزي للعصية Axial hole وتبرز نهايتي الحامض 3 و 5 ممثلتين الذيلين الطويل والقصير من نهاية العصية وتبقيا متدليتان ويبدأ الحامض النووي بالتداخل مع الوحدات البروتينية مرتبطا مع النهايات المرنة للأحماض الامينية في كل وحدة بروتينية ومكونا "الفكة القافلة Lock washer وذلك بتشكل الحامض بطريقة معينة ليكون ثلاث عروات loops، والعروات هي مناطق كثيفة من الازدواج النيوكليوتايدي الداخلي Internal base pairing لجزء من خيط الحامض النووي، وان العروة الأولى المتكونة هي ذات تعاقب AAGAAGUCG وهي أول من يرتبط بالقرص المزدوج لتبتدئ عملية التجميع والذي يمكن أن يحصل بغياب العروتين الثانية والثالثة، إلا أن وجودهما أفضل لإنجاز التجميع. إن تكوين الفكة القافلة عند ابتداء البناء هو افتراض واقعي يفسر الية التجميع ودليل ذلك هو التكون السريع للحلزونات البروتينية العارية بدون حامض نووي عند إجراء خفض سريع للأس الهيدروجيني في تجارب أجريت خارج النسيج الحي، ثم تتكون المزيد من الأقراص المزدوجة التي ترتبط مع القرص المزدوج الأول ليستمر سحب الحامض النووي مع استمرار بناء العصية ويكون سحب الخيط من خلال الثقب المركزي لها وتحديدا للخيط 5 إلى حين اكتمال التجميع ثم يختفي الذيل 5 داخل العصية ويندمج الذيل القصير 3 بالوحدات البروتينية وبذلك يتم بناء الفايروس الجديد.
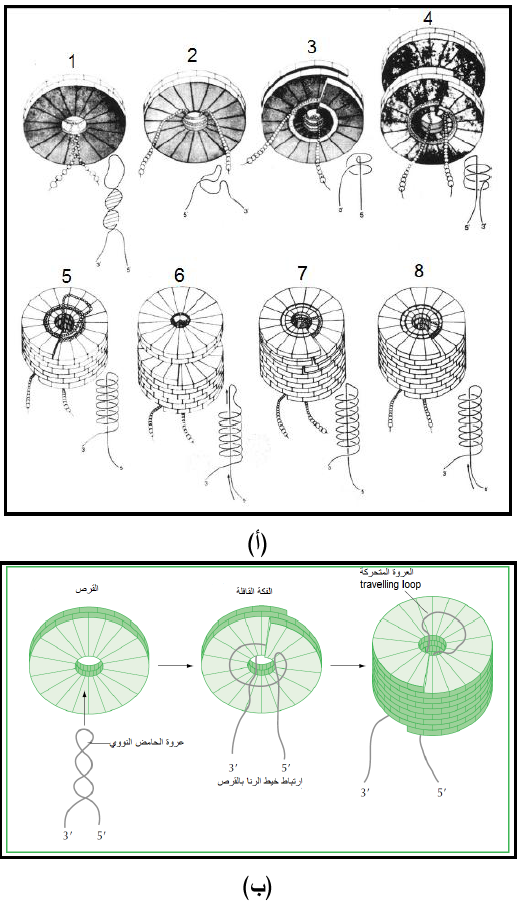
الشكل (8) الية تجميع الغطاء البروتيني مع الحامض النووي لفايروس موزائيك التبغ ممثلا للفايروسات العصوية.
(أ) المراحل الثمانية لعملية التجميع حيث تمثل المراحل من 1-3 عملية الانشاء، وتمثل المراحل من 4-8 مرحلة الاستطالة. وفيما يلي وصف للمراحل الثمانية:
المرحلة (1) تدخل عروة الحامض النووي المسماة "عروة دبوس الشعر Hairpin loop في الثقب المركزي للقرص الاول ذو معامل الترسيب S20 ويكون الدخول من الجانب السفلي للقرص، المرحلة (2) تنفتح العروة للأعلى باستمرار ارتباطها وتشابكها بين طبقتين من الوحدات البروتينية، المرحلة (3) يسبب التداخل بين البروتين والحامض النووي تحويل القرص إلى شكل الفكة القافلة الحلزونية Helical lock washer ليتكون الحلزون الأولي Protohelix ويبرز ذيلي الحامض من نهايته مكونا "معقد الفكة القافلة والرنا " Lock washer-RNA complex وهو بداية تكوين العصية الحلزونية، المرحلة (4) يضاف قرص مزدوج ثاني إلى الأول من الأعلى وتتكون دورتين إضافيتين من الحامض الرايبي بالطريقة ذاتها وكما تبينه المراحل 5-8 وبذلك يستمر بناء العصية الحلزونية باتجاه النهاية 5 للحامض النووي مع استلام عروة الحامض الرايبي للمزيد من الأقراص ثم ينسحب الذيل 5 خلال الثقب المركزي.
(ب) توضيح الآلية تكوين عروة الحامض النووي لفايروس موزائيك التبغ الثلاثية والذيلين 3 و5 وارتباطهما مع القرص المزدوج لتكوين الفكة القافلة ثم استمرار تحرك العروة مع استمرار بناء الأقراص وكما موضح في الشكل الأيمن.
الشكل مقتبس منHull (2002) و Cann ( (2005

الشكل (9): بناء كابسيد فايروس موزائيك التبغ (TMV) من الوحدات البنائية البروتينية بتأثير الأس الهيدروجيني. يتكون الكابسيد من الوحدات البنائية الموحودة Monomeres الملاحظة يمين الشكل والتي تكون حرة مفككة عند قيمة الأس الهيدروجيني 8، ثم تبدأ بالتجمع وبناء الأقراص المزدوجة المرتبطة بنظام الفكة القافلة Lock washer كمرحلة أولية لتجميع الكابسيد عند قيمة 7، ثم يكتمل التركيب الحلزوني للكابسيد عند قيمة 6 ويلاحظ أن عملية التجميع والتفكيك مستمرتين رجعيا بتغير درجة الأس الهيدروجيني.
الشكل مقتبس من Cann (.(2005
4. كيفية بناء الفايروسات الايزومترية (العشرينية الأوجه)
تنتظم الوحدات البنائية البروتينية في كابسيد هذه الفايروسات بشكل أسطح مثلثية Triangular faces والتي تشكل الجسم البلوري الذي ينتظم داخله الحامض النووي بطريقة معقدة بحيث يرتبط مع كل الوحدات البنائية البروتينية في تجويف مغلق داخل البلورة. إن النوع الأكثر شيوعا للبلورات الفايروسية هي البلورات العشرينية الأوجه Icosahedrons وهي البلورة المكونة من عشرين واجهة مثلثية متساوية الأضلاع وفيها 12 سمتا، (الشكل 3) حيث أن كل بلورة لا تبنى بأقل من 60 وحدة بروتينية وذلك لان كل وجه مثلثي مكون على الأقل من ثلاث وحدات بروتينية متماثلة وبوزن جزيئي 50 كيلو دالتون لكل منها. لقد اختير لهذه الفايروسات هذا التركيب المكون من عشرين واجهة مثلثية لأنه الشكل الأكمل إذ يمكن بناء الشكل البلوري من أربعة أوجه مثلثية Tetrahedron أو بشكل مكعب مكون من ستة أوجه رباعية أو من ثمانية أوجه مثلثية Octahedron أو من 12 وجه خماسي Dodecahedron، إلا أن البلورات العشرينية الأوجه هي الأفضل اقتصادا في بناء الجسم وتعطي هيكلا محكما يوفر أفضل حماية للجينوم ويقلص المسافات بين الوحدات البنائية البروتينية إلى الحد الأدنى فإذا ما بني الجسم بشكل رباعي مثلا ستكون الوحدات البنائية البروتينية كبيرة وتصبح المسافات كبيرة بينها مما يقلل من كفاءتها لحماية الجينوم.
عرف الشكل العشريني لأول مرة من قبل Williams و Smith سنة 1958 عند فحصهما للفايروس الحشريTipula iridescent virus استنادا إلى الظل الذي كونه عند وضع جسيماته في مسار الأشعة السينية حيث أعطى ظلا مدببا تارة وآخر مستقيما تارة أخرى عند تدوير الجسم في مسار الأشعة، (الشكل 10).

الشكل (10): استنتاج الشكل البلوري وتخيله بتعريض الجسيمات الفايروسية البلورية للأشعة السينية وملاحظة الظل بعد تدوير الجسيمة حيث ظهر ظلا مدببا ثم مستقيما. الصورة اليسرى هي لجسيمة حقيقية للفايروس الحشري Tipula iridescent Virus،
والصورة اليمنى هي مخطط افتراضي للجسيمة يبدو فيها الظلين المدبب والمستقيم.
الشكل مقتبس من Fraenkel-Conrat (1969).
إن البلورة العشرينية الأوجه هي ابسط أنواع البلورات إلا أن بعض الفايروسات المعقدة التركيب يتكون كابسيدها من أكثر من 60 وحدة بروتينية وقد خلق هذا مشكلة لدى الباحثين في فهم التركيب العشريني لذلك وضع Casper و Klug (1962) نظرية "التكافؤ المشابه "Quasi-equivalence وهي ميل الوحدات المتجاورة غير المتطابقة التجاور للارتباط معا بأواصر متكافئة Equivalent bonds وهذه الحالة لا تحصل إلا في الجسم البلوري العشريني وبذلك فتحت هذه النظرية الباب لفهم كيفية بناء بلورات عشرينية أكثر تعقيدا بنظام شبه عشريني الوجه Near icosahedral packaging وذلك بتكوين مثلثات ثانوية في كل وجه مثلثي رئيسي لتحمل ترتيب المزيد من الوحدات البنائية إذ أن بناء المثلثات الثانوية الإضافية يتطلب مضاعفة الستين وحدة بنائية وهذا يحتاج إلى تطويع شكل هندسي مرن قادر على احتواء هذا العدد المتزايد من الوحدات البنائية وإدخالها بمرونة وإحكام في مواقعها سواء بالطريقة المتكافئة أو شبه المتكافئة وهي عملية صعبة لان الوزن الجزيئي لتلك الوحدات يتراوح بين 30-70 كيلو دالتون ويتم ذلك بتقسيم المثلثات الرئيسية إلى مثلثات ثانوية في النظام البلوري وكما سيوضح ذلك لاحقا. هكذا يتضح أن هناك نوعين من التأصر عند بناء البلورات الفايروسية وهما التاصر المتكافئ Equivalent bonding والذي تكون فيه الأواصر الداخلية متساوية القوة ومتكافئة مما يؤدي إلى انتظام متطابق للوحدات البنائية السداسية المتماثلة في بناء هندسي محكم وهذا ما يحصل في البلورات الفايروسية العشرينية المكونة من 60 وحدة، أما النوع الثاني من التأصر فهو "التأصر شبه المتكافئ" –Quasi equivalent bonding والذي يُظهر تغيرا طفيفا في قوة الأواصر مما يعطي مرونة في ترتيب الوحدات البنائية تسمح بدورها ببناء تراكيب بلورية متنوعة التعقيد، (الشكل 11).

الشكل(11): نوعي التاصر للوحدات البنائية للفايروسات الايزومترية (أ) التأصر المتكافئ ذو الأواصر المحكمة والمكون من كابسوميرات سداسية (ب) التأصر شبه المتكافئ والذي يمتاز بوجود الأواصر الداخلية المرنة بين الوحدات الخماسية المؤشرة بحرف X داخل التركيب الأيسر.
تفهم أشكال الفايروسات الايزومترية بمعرفة أعداد الكابسوميرات السداسية, Pentagonal and hexagonal capsomer, التي يتكون منها الكابسيد وليس هناك الزام ان يكون الجسم البلوري عشريني الوجه بل يمكن بناء البلورة باثنا عشر وجها فتتكون البلورة الإثنا عشرية الوجه Dodecahedral، كما يمكن بناؤها بأكثر من عشرين وجها.
ولفهم بناء هيكل البلورة العشرينية الأوجه السيطة فإن هذا البناء يحتاج على الأقل إلى عشرين مثلث متساوي الأضلاع وكل مثلث يتكون من ثلاث وحدات بنائية متماثلة أو غير متماثلة تنتظم فيه بثلاثة نظم، (الشكل 12)، ولغرض بناء هذه البلورة فان هذا يتطلب استعمال 60 وحدة بنائية على الأقل وهو الشكل الأبسط الذي يكون فيه عدد التثليث= (T) Triangular number) 1 لان الوجه المثلثي بسيط وغير مقسم إلى مثلثات ثانوية، وحيث أن معادلة بناء الهيكل العشريني هي:
عدد الكابسوميرات اللازمة لبناء البلورة = 2 + عدد التثليث X 10
عليه فان عدد الكابسوميرات اللازمة لبناء البلورة وبتطبيق المعادلة المذكورة هي 2 + 1 × 10 = 12 كابسومير، وقد أشير سابقا أن بناء البلورة يتطلب 60 وحدة بنائية على الأقل عليه يجب أن يتكون كل كابسومير من خمسة وحدات بنائية (12 ×5 = 60) وبذلك تتكون ابسط بلورة عشرينية من 12 كابسومير خماسي Pentagon من دون اي كابسومير سداسي Hexagone، إلا انه يمكن تعقيد الهيكل بتقسيم المثلثات الرئيسية إلى مثلثات اصغر متساوية الأضلاع أيضا وبذلك ستصبح هناك حاجة إلى كابسوميرات سداسية إلى جانب الخماسية ويمكن حساب الكابسوميرات السداسية من معرفة عدد التثليث (T) وفق المعادلة التالية:
عدد الكابسوميرات السداسية = (1 - عدد التثليث 10 X) وإذا ما طبقت هذه المعادلة على ابسط بلورة عشرينية ذات عدد تثليث = 1 ، عليه فان عدد الكابسوميرات السداسية يساوي صفر (1-1) × 10 = 0 أي أن هذه البلورة لا تحتاج لأي كابسومير سداسي لبنائها، ولكن بتعقيد الهيكل فان عدد التثليث سيزداد بتقسيم المثلثات الأصلية إلى مثلثات ثانوية اصغر وبذلك نحصل على أعداد تثليث بقيم 1و 4و9 الخ، وكما مبين في الشكل (13) فإذا كان عدد التثليث يساوي واحد فهذا يعني أن الوجه المثلثي مكون من مثلث واحد بسيط وإذا قسم ضلع المثلث إلى قسمين متساويين ستتكون تسع مثلثات فرعية ويمكن أن يزداد تعقيد الجسم إذا ما حدث انبعاج في مركز كل مثلث ثانوي حيث ستتكون مثلثات أخرى تحت فرعية، حيث أن الانبعاج المركزي سيكون ثلاث مثلثات أخرى تحت فرعية (تحت ثانوية) وكما موضح في الشكل (13)، وتحسب أعداد المثلثات التحت ثانوية المتكونة وفق المعادلة التي وضعها ماثيوس وهي : عدد التثليث = عدد المثلثات التحت ثانوية المتكونة نتيجة الانبعاج (هي ثلاثة دائما) × مربع عدد أقسام قاعدة المثلث الرئيسي، وإذا ما طبقت هذه المعادلة على الشكل أعلاه فان أعداد المثلثات التحت ثانوية (عدد التثليث) = 16 وهي، ناتج 3 (عدد المثلثات التحت ثانوية المتكونة نتيجة الانبعاج) × 22 (عدد أقسام قاعدة المثلث الرئيسي)، وهكذا مع أي عدد من أقسام قاعدة المثلث الرئيسي.
إن البناء البلوري الفايروسي هو بناء معقد يتكون مرن التراكيب السداسية Hexamers والخماسية Pentagons واحيانا الثنائية (المثليات) Dimers (1).

الشكل (12): أشكال التجمع المحتملة للوحدات البروتينية في الشكل العشريني البسيط لتكوين الكابسوميرات الممثلة بالأسطح المثلثية حيث تكون كل ثلاث وحدات بروتينية بنائية وجه مثلثي واحد ضمن بناء خماسي.
الشكل مقتبس من Hull ( (2002
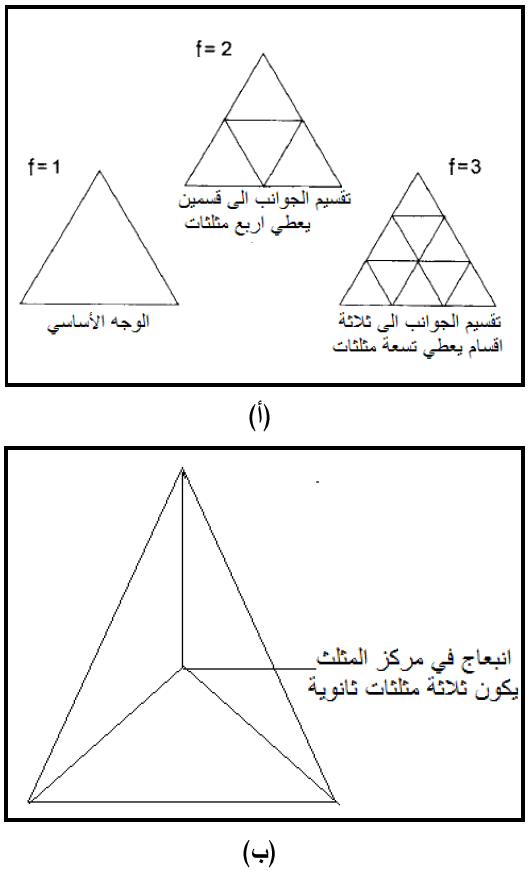
الشكل (13): طريقة تقسيم الأوجه المثلثية الرئيسية إلى مثلثات ثانوية Sub-triangles عند بناء النظام البلوري الفايروسي المعقد.
(أ) تقسيم الوجه المثلثي الأساسي إلى مثلثات ثانوية بتقسيم قاعدته إلى أقسام متساوية، لإعطاء مثلثات ثانوية، فعندما تكون قاعدة المثلث بسيطة غير مقسمة يتكون مثلث واحد بسيط، وعندما تقسم قاعدة المثلث إلى قسمين تتكون أربعة مثلثات أي يصبح عدد التثليث 4، وعندما تقسم قاعدة المثلث إلى ثلاثة أقسام تتكون تسعة مثلثات وهكذا. تشير الأرقام أسفل كل مثلث إلى عدد أقسام قاعدة المثلث. والقاعدة التي تحكم ذلك هي: عدد المثلثات الثانوية المتكونة (عدد التثليث) = (عدد أقسام قاعدة المثلث)2، فيما تشير قيمة f إلى عدد اقسام ضلع المثلث أي عدد الاقسام الثانوية لكل جانب من الوجه المثلثي.
(ب) تقسيم المثلث إلى مثلثات ثانوية نتيجة انبعاج في مركز المثلث.
الية تجميع الفايروسات الايزومترية
لا يعرف الكثير عن آلية تجميع هذه الفايروسات ولكن أقترح Matthews (1991) الية لتجميع فايروس الموزائيك الاصفر للشلغم (TYMV) ممثلا لهذه الفايروسات والذي يتضاعف في الكلوروبلاستات وكما مبين في الشكل (14) الذي يظهر تخليق الكابسوميرات الخماسية والسداسية بواسطة الشبكة الاندوبلازمية وتراكمها في السايتوبلازم محيطة بالحويصلة الموجودة في الكلوروبلاست، ثم تدخل هذه الكابسوميرات في الغشاء الخارجي للكلوروبلاست وتنطمر بالغشاء الليبيدي المزدوج بارتباط جوانبها الكارهة للماء Hydrophobic Sides معا داخل الكابسيد المنطمر هو الأخر في الغشاء الليبيدي، ويتم تخليق الرنا داخل حويصله الكلوروبلاست ثم يبدأ بالعبور خلال عنق الحويصلة حيث يتم في هذه المرحلة التعرف على تعاقب نيوكليوتايدي معين فيه ويحدث الارتباط مع الكابسوميرات الخماسية والسداسية أيضا وذلك من مصدر التجهيز وهو غشاء الكلوروبلاست وبعدها تطلق الجسيمات الفايروسية إلى السايتوبلازم.

الشكل (14): آلية تجميع فايروس الموزائيك الأصفر للشلغم (TYMV). يبين الشكل أن الحامض النووي للفايروس والمخلق داخل حويصلة الكلوروبلاستيدة يكتسب الكابسوميرات الخماسية (الخماسيات Pentagones) و الكابسوميرات السداسية (السداسيات Hexamers) المتكونة في غشاء الكلوروبلاستيدة عند عبوره عنقها متوجها إلى السايتوبلازم.
الشكل مقتبس من Hull (2002)
5. كيفية بناء الفايروسات المغلفة
يبنى نيوكليوكابسيد هذه الفايروسات بذات الأليات التي ذكرت لكل من الفايروسات العصوية والأيزومترية وذلك لأن شكل نيوكليوكابسيدها هو إما عصوي أو أيزومتري وحسب نوع الفايروس المغلف وبالتالي فإن هيكليتها تماثل هيكلية الفايروسات المذكورة، إلا أن الفرق المهم في تركيب هذه الفايروسات هو وجود الغلاف الليبوبروتيني والكلايكوبروتيني الذي يحيط بالنيوكليوكاسيد والذي يعقد تركيب هذه الفايروسات، وليس بالضرورة أن يكون شكل الغلاف مماثلا لشكل النيوكليوكابسيد فقد يكون الأخير عصويا أو بلوريا والغلاف كرويا وكما مبين في الشكل (15) الذي يظهر مخططا لمقطع للتركيب العام لفايروسين مغلفين من فايروسات الرابدو Rhabdoviruses والتوسبو Tospoviruses.

الشكل (15): الفايروسات المغلفة (أ) جسيمة مغلفة عصوية معقدة تمثل فايروسات الرابدو Rhabdoviridae مصورة بالمجهر الالكتروني (ب) جسيمة مغلفة كروية معقدة مصورة بالمجهر الالكتروني (ج) مخطط لمقطع في جسيمة لفايروس رابدو المغلفة العصوية (د) مخطط لمقطع في جسيمة لفايروس من فايروسات التوسبو Tospoviruses المغلفة الكروية.
الشكل مقتبس من Mahy و (2008) Van Regenmortel وCann (2005).
 الاكثر قراءة في الفايروسات والامراض التي تسببها للنبات
الاكثر قراءة في الفايروسات والامراض التي تسببها للنبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















