























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Origin of Eukaryotic Cells
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 19-11-2016
19-11-2016
 2709
2709




+
-
20
Origin of Eukaryotic Cells
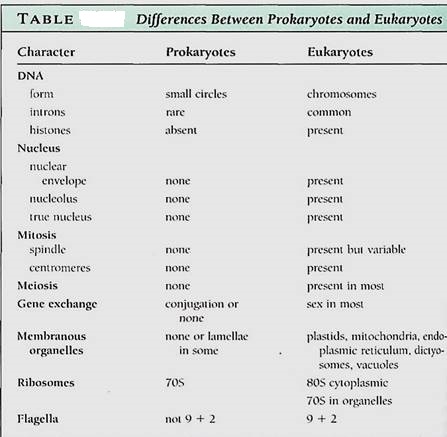
The differences between prokaryotes and eukaryotes are deep-seated, involving many of the most basic metabolic processes and cellular organization (Table ).
DNA STRUCTURE
In prokaryotes, DNA is "naked," not complexes with any proteins, its numerous negative charges being neutralized by calcium ions instead. Each genome is just a short circle of DNA, often containing only about 3000 genes. The DNA of prokaryotes contains few introns, and processing of mRNA is relatively simple.
In eukaryotes, DNA is more elaborately packaged, being complexes with the nucleosome histones and forming a chromosome . Each molecule of DNA is long, capable of carrying thousands of genes, yet no genome ever consists of just a single DA molecule; at least two occur per haploid set, and 10 to 20 are more common. Many eukaryotic genes contain introns, and large segments of DNA do not code for any type of RNA.
NUCLEAR STRUCTURE
The DNA of prokaryotes lies directly in the cytoplasm; no nucleus is present. Most of the DNA of eukaryotes is located within a nucleus, separated from the cytoplasm by two nuclear membranes with nuclear pores; this is the source of their name: eu- (true) karyotes. A nucleolus is present as well.
NUCLEAR DIVISION
As a prokaryotic cell grows, its plasma membrane expands; the DNA circles attached to it are separated from each other by cell growth. The cell then pinches in two.
Eukaryotic cells are typically haploid or diploid, having only one or two copies of each of their various chromosomes, so the rather haphazard prokaryotic method of simple membrane expansion would not work reliably. Instead, mitosis has evolved in eukaryotes: Spindle microtubules, cell plate formation, and centromeres ensure that each progeny cell receives one of each type of chromosome . In almost all eukaryotes, meiotic nuclear division occurs as part of sexual reproduction. The critical phenomena are pairing of maternal and paternal homologous chromosomes followed by crossing-over and genetic recombination. The complex procedures result in an interchange of alleles, such that a great diversity of spores or gametes is produced.
ORGANELLES
Prokaryotes lack membrane-bounded organelles: Their cytoplasm is rather homogeneous, containing only ribosomes and perhaps gas vacuoles, storage granules, or photosynthetic membranes, depending on the species. Eukaryotes all have nuclei and mitochondria, and plants and algae also have plastids. In addition, dictyosomes, endoplasmic reticulum, vacuoles, and vesicles are all parts of the eukaryotic endomembrane system. These membranes result in the division of the cell into numerous compartments, permitting several types of highly specialized metabolism to occur within the same cell. The ribosomes of prokaryotes are 70S, being smaller and denser than the 80S ribosomes in the cytoplasm of eukaryotes . Plastids and mitochondria have 70S ribosomes instead of 80S ribosomes.
Flagella and cilia are remarkably uniform in eukaryotes, consisting of a 9 + 2 arrangement of microtubules. Only a few prokaryotes have flagella, and these have a totally different type of construction, never a 9 + 2 arrangement. They are not composed of microtubules or tubulin.
Numerous other distinctions could be mentioned, especially in gene regulation and biochemical pathways. The differences are numerous and basic, much more so than the differences that distinguish plants from animals.
ORIGIN OF EUKARYOTES: THE ENDOSYMBIONT THEORY
Prokaryotes represent the types of organisms that arose first. Until the early 1970s, it was assumed that some had given rise to eukaryotes by gradually becoming more complex and developing an endomembrane system with mitochondria and plastids. This is the autogenous theory. Cyanobacteria, then known as blue-green algae, seemed to be logical inter-mediates
However, in the 1960s the endosymbiont theory was revived. It had been proposed in 1905 by K. C. Mereschkowsky, who had speculated that plastids and mitochondria might be prokaryotes living inside eukaryotic cells. At that time there was no way to test the theory, but by the late 1960s, plastids and mitochondria were discovered to have their own DNA and ribosomes that are 70S, not 80S. Furthermore, these organelles divide by a type of cleavages similar to that of prokaryotes, and they also lack microtubules. Before long, other prokaryotic traits were discovered in them: Their DNA is a small closed circle of naked DNA, and organellar genes are organized like prokaryotic genes. Their ribosomes are sensitive to the same antibiotics that interfere with prokaryotic ribosomes.
The endosymbiont theory postulates that a prokaryotic cell had evolved to the point of having some eukaryotic features such as 80S ribosomes and a nuclear envelope (Fig. 1). It was postulated to be heterotrophic, living by engulfing and digesting other cells. It may have engulfed one but then not digested it, perhaps because the larger cell had inefficient digestive enzymes or because the engulfed cell had a more resistant membrane. Such an association might be advantageous: The engulfed cell could be extremely efficient at aerobic respiration but poor at obtaining nutrients. If it could get sugars from the larger cell and "leak" ATP back to it, both cells might be better off. An endosymbiotic relationship might be more advantageous selectively than the condition in which each partner is living independently. Such an endosymbiosis would still have many obstacles to overcome— increasing the compatibility between the two cells and coordinating growth, development, and reproduction—but the descendants of the engulfed organism could possibly evolve into a mitochondrion.
Origin of Plastids. Plastids could arise similarly if the engulfed partner were photosynthetic (Fig. 1). Because all chloroplasts contain chlorophyll a and produce oxygen but none have bacteriochlorophylls, we suspect cyanobacteria as possible ancestors and do not believe that the green and purple bacteria are very likely candidates. Prochlorophytes arouse great interest because they have both chlorophyll a and b and some lack phycobilin pigments . The first prochlorophyte discovered, Prochloron, not only resembles chloroplasts biochemically but also exists as an obligate symbiont inside marine invertebrates, called didemnid ascidians, supplying them with carbohydrates whenever the ascidians lie in the sunny photic zone. Prochloron has never been isolated and cultured outside the tissues of the ascidian, so research with it has been difficult; however, a second prochlorophyte, Prochlorothrix, has been discovered growing freely in lakes in Holland. 16S rRNA sequencing has now shown that chloroplasts, Prochlorothrix, and Prochloron are all definitely closely related, but neither bacterium is the immediate ancestor of chloroplasts. Instead, they all share a common ancestor.
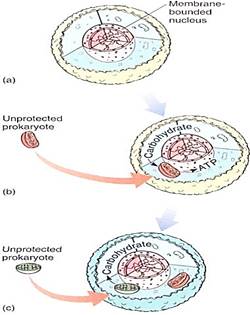
FIGURE 1:The endosymbiont theory postulates that a prokaryote (a) evolved to the level of having a membrane-bounded nucleus; this must have been an early step, because all eukaryotes have a nuclear envelope, and all are rather uniform in structure. (b) The early eukaryote engulfed but did not digest a prokaryote, which then evolved into a mitochondrion. (c) Finally, the descendants of that early eukaryote would engulf but not digest a prochlorophyte type of cyanobacterium, which would evolve into a plastid; this would have been the point of separation of the animal and plant lines of evolution. Not shown in the diagram is the evolution of mitosis, meiosis, sexual reproduction, and cell walls.
How many steps of endosymbiosis might have occurred? At least two, one for mitochondria and a second lor plastids. Because all eukaryotes have mitochondria and only some have plastids, we suspect that mitochondria arose first.
Chloroplasts of red algae may have arisen by an act of endosymbiosis distinct fom the one that produced the plastids of green algae. Red algal chloroplasts are remarkably cyanobacteria in organization and biochemistry, having phycobiliproteins and phycobilisomes arranged on membranes that do not form grana stacks (Fig. 2a). They store a polysaccharide, floridean starch, that resembles glycogen.
Although it is possible to construct a hypothetical pathway by which a red ^ chloroplast could evolve into a green algal chloroplast, such a hypothesis has many difficult and highly unlikely steps. Also, it would mean that red algae would have been ancestral to green algae, and many cell characteristics are not compatible with such a conclusion. It is more plausible that red algae represent a distinct line of evolution from the early eukaryotes, its chloroplasts being evolutionarily distinct from the chloroplasts of green algae and land plants.
Brown algae may represent a third line of chloroplast evolution (Fig. 2b). Brown algae, along with golden brown algae, diatoms, and yellow-green algae (all three in division Chrysophyta) and dinoflagellates (division Pyrrhophyta) all contain chlorophyll c in addition to chlorophyll a, and they all lack chlorophyll b. Their non-chlorophyll accessory pigments are also unique, being types of carotenoids not found in green algae or plants. Membranes of brown algal chloroplasts associate into grana-like stacks, but they are always small, each consisting of just two or three membranes.
If there really were three or four separate prokaryotic ancestors to three or four distinct lines of chloroplasts, the plastid DNA nucleotide sequences of each line should show little homology to the others. But if one type evolved from another—green algae evolving from brown algae for instance, considerable sequence homology is expected. Studies are currently underway, and results should be known soon.
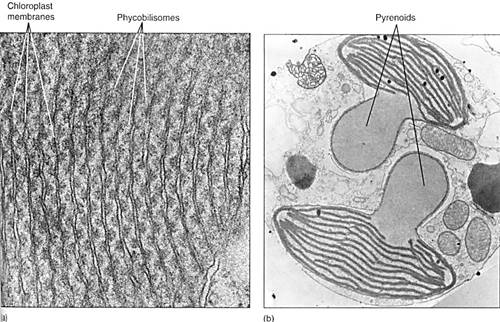
FIGURE 2: (a) Internal membranes of red algal chloroplasts do not form grana stacks like those of green plants. Their phycobilisomes are visible, attached to the membrane. Porphyridium (X 115,000). Curesy of E. Gantt and S. F. Conti, University of Maryland) (b) Brown algal chloroplast membranes associate into stacks of three or, less often, stacks of two. The large clear structures are pyrenoids, involved in polymerization of sugars into polysaccharides.
Origin of Nuclei. The nuclei of plants are virtually identical to those of animals in terms of structure, metabolism, mitosis, and meiosis. Apparently the plant and animal lines of evolution diverged only after the eukaryotic nucleus had become quite sophisticated. But nuclei are complex organelles, and mitosis and meiosis are intricate processes with many steps. Certainly plant and animal nuclei could not have arisen from prokaryotic nucleoids quickly; many intermediate steps spanning hundreds of millions of years must have been involved.
Several groups of organisms with unusual nuclear characteristics may represent lines of evolution that originated earlier in the history of eukaryotes than did plants and animals. the nuclei of many fungi were shown to have an unusual mitosis, some having both an intranuclear and an extranuclear spindle. In many fungal species the nuclear envelope and nucleolus either do not break down at all or do so only late in mitosis. Similar behavior occurs in many groups of algae, including dinoflagellates, brown algae, euglenoids, and some green algae (Fig. 3a). Gaps form in the nuclear envelope, and bundles of microtubules pass completely through the nucleus.
Even more significantly, nuclei of dinoflagellates have no histones. Thy do have what appear to be chromosomes, but these remain condensed at all times, undergoing only a slight uncoiling during interphase (Fig. 3b and c). Because histone and nucleosome structure is so constant in all other eukaryotes, this unique situation in dinoflagellates is considered highly significant. It has been proposed that dinoflagellates be removed from eukaryotes and classified as mesokaryotes (meso, intermediate). Dinoflagellates are receiving a great deal of attention.
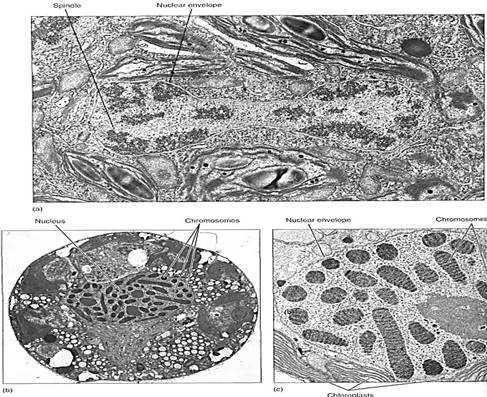
FIGURE 3: (a) Late telophase in the green alga Cladophora; not only is the nuclear membrane completely intact, but a portion of the intranuclear spindle is visible (X 26,656). (Courtesy of J. L. Scott and K. W. Bullock, College of William and Mary) (b) An interphase nucleus of Peridinium, a dinoflagellate; chromosomes never completely decondense. It is not known how chemical messengers, DNA replicases, and RNA synthetases gain access to the DNA molecule (X 14,000). (Courtesy of J. M. Chesnick, University of Washington) (c) Magnification of dinoflagellate chromosomes (X 21,000). (Courtesy of J. Dodge, University of London).
Types of Cytokinesis. Several types of cytokinesis occur in algae. Many groups of unicellular algae have no wall, their plasma membrane being their outermost surface. During cell division, the plasma membrane pinches in two, being pulled inward as a cleavage furrow, a process remarkably similar to cytokinesis in animals (Fig. 4a).
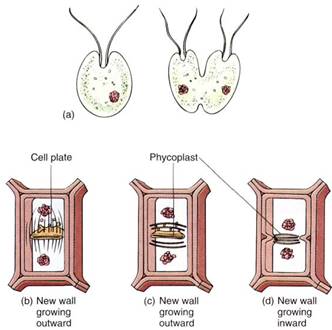
FIGURE 4: (a) In unicellular, wall-less forms, cell division occurs by infurrowing of the plasma membrane. (b) Most algae with walls divide by a phragmoplast and cell plate, just as plant cells do. (c) and (d) Some green algae have a phycoplast; the mitotic spindle disorganizes quickly, after which a new set of microtubules polymerizes perpendicular to the spindle axis. Division may then occur by cell plate growth (c) or by infurrowing (d).
In almost all algae with walls, cell division is similar to that of plants: After telophase, He spindle microtubules persist temporarily, holding the nuclei apart. Between them a phragmoplast forms, consisting of short microtubules oriented parallel to the spindle microtubules. The coalescence of dictyosome vesicles establishes a cell plate, which then grows radially outward—centrifugal growth—until it meets and fuses with the wall of the parental cell.
In some green algae, cytokinesis occurs by a different method; the mitotic spindle depolymerizes quickly, and the two daughter nuclei lie close together. A new set of microtubules appears between them, oriented parallel to the plane where the new wall will form, which is perpendicular to the orientation of the spindle. This group of microtubules is a phycoplast and may be associated with division either by furrowing or by cell plate formation (see Fig. 4c and d). Phycoplasts seem to be associated with those green algae whose nuclear envelope does not break down during mitosis, whereas phragmoplasts are associated with the loss of the nuclear envelope. Plants probably arose from green algae that divide with a phragmoplast rather than a phycoplast.
In red algae, cell division occurs by a phragmoplast, but the new wall grows inward from the pre-existing walls. It stops growing while still incomplete, having a large hole called a pit connection, which is filled with material (a pit plug; Fig. 5). Otherwise, most aspects of wall formation are similar to those of other walled algae.
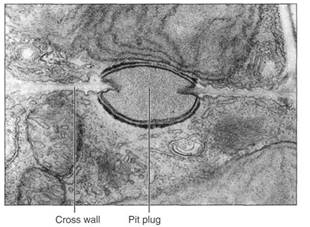
FIGURE 5:Cross walls of red algae are unusual in being incomplete, having a hole in their center. This is closed by a pit plug. Scinaia conjusa (X 15,000). (Courtesy of Curl Pueschell, State University of New Yarn Binghamton)
Origin of Flagella. The origin of flagella is still unknown. Being composed of microtubules, flagella may be related in origin to the mitotic spindle. Flagella must have occurred extremely early because all flagella and cilia of eukaryotes have virtually identical internal structure. The standard composition is an external membrane that is an extension of the plasma membrane and an internal axoneme composed of nine outer doublets and two inner singlet microtubules . Because the structure is so constant, we are quite confident in excluding the possibility of multiple origins and convergent evolution. Because virtually all eukaryotes have flagella or cilia, this structure must have arisen Ife the various lines diverged.
Variability exists in the external covering and length of flagella (Fig. 6). Many algal and fungal and all plant flagella have a smooth surface and are known as whiplash flagella. Others have a surface covered with hairs and are referred to as fad flagella. Flairs are of multiple types; some appear to be just a single row of protein monomers, whereas others are hollow tubes of protein that end as a single row.
Considerable attention is being given to both the tips and the bases of flagella, although studies of both are difficult. The central microtubules extend beyond the peripheral ones, and the arrangement at the tip varies from group to group. In many species, the central microtubules extend a long distance beyond, forming an extremely fine hair tip that is an adhesive recognition organelle. In gametes, the hair tips of one gamete recognize and bind to those of a compatible gamete. Once the flagellar tips adhere, the gametes are drawn together and fusion of plasma membranes and cytoplasm occurs. In some swimming spores (zoospores), flagellar hair tips appear to recognize suitable substrates; the connect substrate triggers adhesion, and the spore then settles and germinates. If the flagellum encounters an unsuitable substrate, adhesion and germination do not occur.
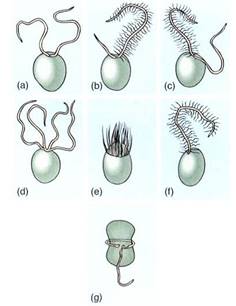
FIGURE 6 :The most common arrangements of flagella in algae. (a) Two equal whiplash. (b) A short whiplash and a long tinsel. (c) Two equally long flagella, one of each type. (d) Four whiplash. (e) Numerous flagella located in a ring (rare). (f) One tinsel flagellum (mostly just in some brown algae). (g) One flagellum trailing, one in a transverse groove (dinoflagellates).
The base of a flagellum is attached to a basal body, which is part of a large, complex root structure. The root structure of eukaryotic flagella is an extensive, elaborate complex of bracing structures that interconnect the basal body and flagellum to arrays of microtubule extending deep into the cell (Fig. 7). Analyzing root structure organization is extremely difficult because they are much too small to be seen by light microscopy yet much too large to extract whole. Serial sections must be cut extremely carefully and every section examined by transmission electron microscopy; from the numerous micrographs, the structure can be reconstructed. Despite the difficulty, phycologists have been successful analyzing many root structures, which are useful in understanding not only the workings of flagella, but also flagellar growth, duplication, and evolution.
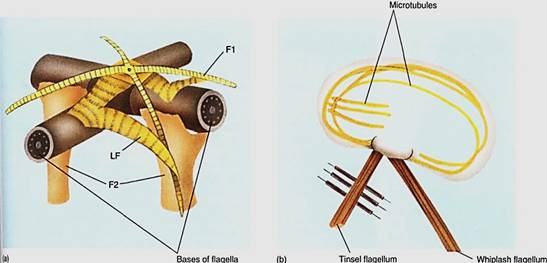
FIGURE 7 :(a) Model of the root structure of Urospora, a green alga, with two massive root fibers (F2) and four narrower ones (F1). All are interconnected and bound to the flagella by lateral fibers (LF). (b) The root structure of brown algae is anchored to the cell by long bands of microtubules. In this generalized diagram, some microtubules extend the entire cell length, and others extend about one third the length.
Many but not all motile algae have eyespots, organelles involved in phototaxis (swimming toward or away from light). The structure of eyespots is variable: All contain at least globules of lipid, but some also contain carotenoids dissolved in the lipid, and many are associated with specialized membranes. Whereas eyespots are of special importance to photosynthetic organisms, motile gametes such as sperms swim guided by chemotaxis. The chemoreceptors are not at all well understood, although in many species, the attractant sex hormones have been discovered.
Eukaryotes evolved gradually in many steps, and we have numerous organisms that perhaps retain some intermediate, transitional features. Although we do not know how all HE organisms are interrelated, we are certain that no single line of eukaryotes at some point diverged into plants, animals, and fungi. Rather, many protistan groups represent distinct phases in the evolution of eukaryotes. Eventually, the artificial kingdom Protista may be replaced by three, four, or more natural kingdoms.
The divisions of algae that contain predominantly unicellular individuals are discussed below first because they are the simplest and many seem to be relictual, perhaps having originated early. The three groups that contain predominantly multicellular individuals are ten considered.
 الاكثر قراءة في الطحالب
الاكثر قراءة في الطحالب
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















