Division Chlorophyta: Green Algae
From an evolutionary standpoint, the green algae (Table) constitute an extremely important group: Not only could some early green algae organize complex differentiated multicellular bodies, but some moved onto land—the ancestors of true plants, kingdom Plantae. Perhaps because most of the animals and plants with which we are familiar are multicellular and terrestrial, we tend not to appreciate just how complex these phenomena are. Consider that organisms spent as many as 1 billion years at the level of unicellular organization; then among algae true multicellularity evolved only a few times. The transition to living on land was so complex that it occurred no more than two or three times at most, perhaps only once.
Green algae have remarkable developmental and metabolic plasticity: They are resilient and survive many types of disturbances and changes. Mutations that cause cells to adhere after cytokinesis have not been lethal, nor were many mutations that affected swimming, orientation of cell division, or coordination of karyokinesis and cytokinesis. Bodies composed of several types of specialized cells have not evolved in most other algal groups. Although brown algae and red algae do show considerable sophistication in certain types of multicellular bodies, they are metabolically intolerant of ecological changes; few can live in fresh water, soil, air, or inside animals as many green algae do.
The diversity in the green algae is tremendous, but some of the most important evolutionary lines and developments are involved. These give you a good basis for understanding the diversity possible among living organisms and appreciating that the metabolisms and organizations of humans and flowering plants are by no means the only solutions to biological problems.

BODY CONSTRUCTION IN THE GREEN ALGAE
The most relictual and simple body is probably the motile single cell. Numerous green algae are unicellular. From this, many evolutionary possibilities exist:
1. Motile colonies (Figs. 1a, 9, and 10): If cells adhere loosely, the resulting structure is a colony, not an individual organism. All cells are similar and none is particularly specialized; there may be some differentiation into two or three somewhat distinct cell types.
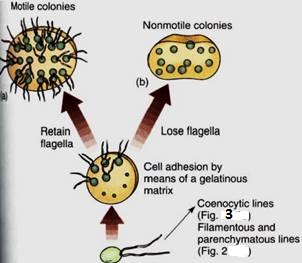
FIGURE 1:Evolution of colonial body type in green algae. The association of cells is controlled, not random. Depending on the species, each colony has a specific number of cells held in a particular shape such as a sphere, a flat plate, or a curved plate. No cell is free to divide at random.
2. Non-motile colonies occur if the cells lose their flagella or never develop them (Fig; 1b). Although a non-motile cell is simpler than one with flagella, the flagellated form is believed to be ancestral because all motile algae have the same type of flagella the 9 + 2 arrangement of microtubules.
3. A filamentous body results if cells are held tightly by a middle lamella and if all cells divide transversely (Figs. 2a and b and 11). If occasional cells undergo longitudinal division, the filament branches. Often some portions of the body differ from others; for example, one end may serve as a means to attach the filament to a lock (a holdfast), or other parts may produce spores or gametes.
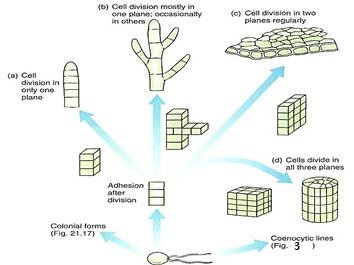
FIGURE 2:Evolution of filamentous, membranous, and parenchymatous body types in green algae. The formation of a multicellular body requires control over timing, position, and orientation of cell division, along with cell-cell adhesion mediated by a middle lamella. (a) All cells divide in only one plane, resulting in a uniseriate filament. (b) If cells occasionally divide in a different plane, the filament branches. A morphogenic mechanism may signal which cells can divide to branch and which cannot. (c) A two-dimensional sheet of cells results if all cells divide in two dimensions regularly. (d) A three dimensional body is produced if cells divide regularly in all three planes. All cells share plasmodesmata with all adjacent cells because all walls result from a cell division cell plate. This is a true parenchyma tissue.
4. A membranous body results if the orientation of cell divisions is controlled precisely such that all new walls occur in only two planes (Figs. 2c and 12). The result is a sheet of cells that can become more extensive but remains thin. A membranous body is more strongly affected by currents and wave action and more likely to be tm than is a filamentous body.
5. If cell division occurs regularly in all three planes, a bulky, three-dimensional parenchymatous body results (Figs. 2d and 21.31). All cells are interconnected by plasmodesmata, and a true parenchyma tissue is formed.
6. A coenocytic or siphonous body results if karyokinesis occurs without cytokinesis, and giant multinucleate cells result (Figs. 3 and 14). The cells can grow to several centimeters in diameter, but they usually remain fairly simple despite their size. Some coenocytes, such as Acetabularia, can be surprisingly complex (Fig. 4):
Evolutionary development of control over the orientation of cell division and adhesion must have been difficult. The fossil record indicates that the ability to grow as a siphonous form came into existence 230 million years earlier than the ability to organize even the simplest filament.
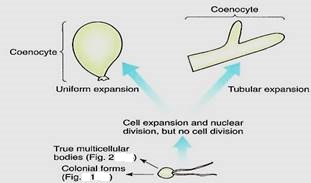
FIGURE 3 :Coenocytic bodies in green algae may be more or less isodiametric (spherical) or long, branched or unbranched tubes. Tubular forms are called siphonous forms.

FIGURE 4:Individuals of Acetabularia are unusual for being giant single cells, but even more remarkable, they are uninucleate. Each cap is composed of hollow tubes of cytoplasm and vacuole, and at times of reproduction, the sole nucleus begins rapid, repeated mitosis, resulting in hundreds or thousands of nuclei. Cytoplasmic streaming carries them upward into the cap segments; when each segment has several hundred nuclei, it either becomes a dormant cyst and is released or produces hundreds of gametes. (Dr. DennisKunkel/Photake).
LIFE CYCLES OF THE GREEN ALGAE
The angiosperm type of life cycle, an alternation of heteromorphic generations, can be traced to the green algae, and the theory of life cycles can be discussed here. In unicellular algae that have no sexual reproduction, such as euglenoids, the life cycle is really just the cell cycle; mitosis and cytokinesis constitute reproduction. But with the evolution of sex, two processes are critical: meiosis, which segregates out sets of chromosomes, and syngamy, which brings two sets back together.
After sexual reproduction evolved, while all organisms were still unicellular (no multi- cellular organism is relictually asexual), the simplest life cycle may have been the following: A diploid cell undergoes meiosis, and each cell then exists as a haploid individual, able to undergo mitosis. Because this is a unicellular organism, cell division is both "growth" and asexual reproduction. Some of these cells can also act as gametes and fuse, producing a diploid zygote that also is able to reproduce asexually—and simultaneously grow—by mitosis. Finally, at some time some of these undergo meiosis again. Little difference exists between gametes, zygotes, and individuals, none of which is very specialized. Both haploid and diploid cells are capable of growth, division, and reproduction. Such species are dibiontic; that is, there is an alternation of generations between haploid and diploid.
In monobiontic species, specialization occurs in that only one free-living generation exists. In some monobiontic species the haploid phase represents the individual and the only diploid cell is the zygote, which is capable only of meiosis, not growth or mitosis (Fig 5a and b). The haploid body, either uni- or multicellular, carries out photosynthesis and growth. In other monobiontic species, the diploid phase represents the individual vegetative growth phase; the only haploid cells are the gametes, which can undergo only syngamy (Fig. 5c and d).
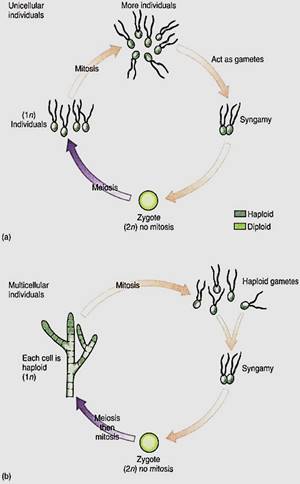
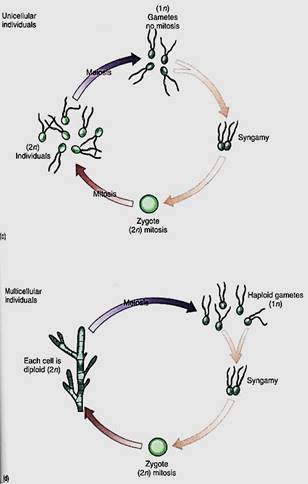
FIGURE 5:In monobiontic life cycles, only one generation—one phase—is capable of undergoing mitosis. In most algae (a and b), the haploid phase is dominant, undergoing mitosis that results in either more unicellular individuals or a multicellular individual. Only one diploid cell occurs—the zygote—and it cannot undergo mitosis; it cannot produce more diploid cells, (c and d, facing page) In a few algae and in all animals, the monobiontic life cycle is dominated by the diploid phase, the only haploid cells being the gametes.
In a dibiontic species in which both stages are multicellular, the gametophyte (haploid phases) and sporophyte (diploid phase) may resemble each other strongly, and alternation of isomorphic generations occurs (Fig. 6a). Or the two may be very different in appearance and construction (alternation of heteromorphic generations), which allows tarn to exploit different ecological niches almost as if they were two species, and gametophytes do not compete directly with sporophytes (Fig. 6b).
All sporophytes, both in algae and in embryophytes, produce spores by meiosis, but may algal sporophytes also produce spores by mitosis; these are diploid and grow into a new sporophyte in a form of asexual reproduction. Some algal gametophytes produce spores by mitosis; these are haploid and develop into new gametophytes, also a form of asexual reproduction.
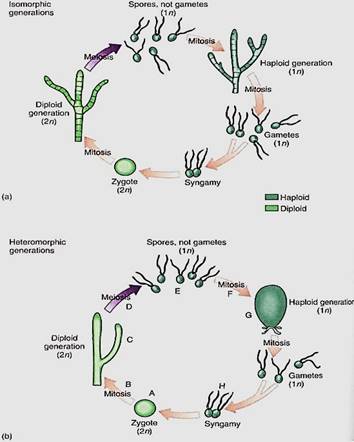
FIGURE 6:Dibiontic life cycles involve an alternation of either isomorphic (a) or heteromorphic (b) generations. With isomorphic generations, the similarity of body types can be so great that is very difficult to know if an individual is a gametophyte or a sporophyte. (b) In dibiontic algae with an alternation of heteromorphic generations, the two types of individuals could never be confused. In flowering plants, the mitosis at B would be the growth of the embryo into the sporophyte plant body, such as an oak tree or a lily, and the meiosis at D would occur only in sporogenous cells of anthers and ovules. The mitosis at F would be the formation of the tube nucleus, generative nucleus, and sperm cells in pollen and the growth of the megaspore into the antipodals, central cell, synergids, and egg cell of the megagametophyte. The syngamy at H would be fusion of one sperm nucleus with the egg. In flowering plants, the zygote begins growth immediately without a resting period, and of course flagella are not formed at any stage.
During the earliest stages of the evolution of sex, the gametes were tsogamous (identical) but anisogamy (slight differences in gametes) and oogamy later evolved (Fig. 7). The reproductive tissues and organs of different algal, fungal, and plant groups differ markedly. At one time specialized terms were applied to every group, resulting in an overwhelming nomenclature. It has since been standardized, so that regardless of the group, gametes are produced in gametangia. Sperm cells or microgametes form in micro- gametangia, and egg cells or megagametes form in megagametangia. Spores are formed in sporangia, either megasporangia or microsporangia, depending on the size of the spore.
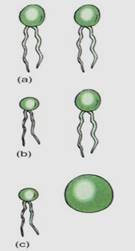
FIGURE 7: (a) Isogametes are identical gametes. Theoretically they could be either motile or nonmotile, but in reality, all known isogametes do have flagella. (b) If the gametes have any visible differences but are still similar, they are anisogametes. (c) Oogametes are obviously different in size, and the megagamete is virtually always nonmotile. In most algae and plants the microgamete has flagella. In flowering plants, conifers, and some algae, microgametes are also nonmotile.
REPRESENTATIVE GENERA OF THE GREEN ALGAE
Unicellular Species. Chlamydomonas is one of the simplest chlorophytes. It is unicellular and, like all green algae, has chlorophyll a and b, carotenoids, and xanthophylls; its starch is formed in chloroplasts just like that of true plants (Fig. 8). It has two anterior flagella, like most motile green algae, and it has normal mitosis, meiosis, and syngamy. Its life cycle is simple: A haploid cell resorbs its flagella, divides mitotically, and forms 2, 4, 8, or 16 new cells that grow new flagella, each of which swims until it encounters a compatible cell. The cells recognize each other by reactions at the tips of their flagella. Hey undergo plasmogamy and karyogamy, forming a large zygote that sheds the four flagella and sinks to the bottom in a quiescent state. Germination is by meiosis to form for biflagellate haploid individuals; the zygote is the only diploid cell.
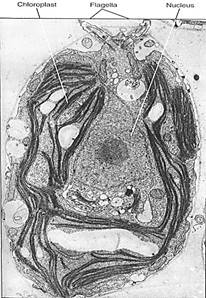
FIGURE 8:The unicellular, haploid green alga Chlamydomonas. Its nucleus, chloroplast, and pyrenoid are visible, but only the bases of the two flagella can be seen. The flagella are at the anterior end; that is, they pull the cell forward rather than pushing it. Several starch grains are present. (U. Goodenough, Washington University).
Motile Colonial Species. In the motile colonial line of evolution, cells that greatly resemble Chlamydomonas are produced when the zygote divides, but the progeny cells are held together by a gelatinous matrix (Fig. 9). In Gonium, each colony contains only a few cells (4, 8, 16, or 32), and the only sign of organization is that all flagella beat in a coordinated fashion. Pandorina is about the same size as Gonium but is slightly more derived because it shows a trace of differentiation; the colony swims in one direction, and anterior cells are slightly different from posterior ones. Volvox is the stunning conclusion of this line: Its colonies can contain up to 50,000 Chlamydomonas-like cells and are easily visible without a microscope (Fig. 10). Differentiation exists in that up to 50 cells in the posterior half of a colony are specialized for reproduction only.
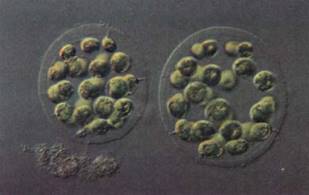
FIGURE 9:Eudorina elegans is a small motile colony of green algae. In each species, the number of cells and their spatial arrangements are constant. The numbers and orientations of cell divisions are controlled. (Walker/Photo Researchers).
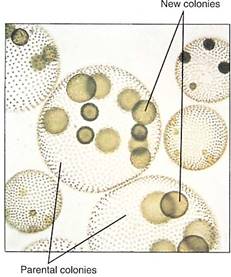
FIGURE 10:Individuals of Volvox are giant spherical colonies containing up to 50,000 cells. The colonies are easily visible to the naked eye. (Manfred Kage/Peter Arnold, Inc.).
Filamentous Species. Members of the genus Ulothrix are simple types of filamentous green algae (Fig. 11a). Their life cycle is monobiontic; it has only one free-living multicellular generation, and it is haploid. It consists of one row of cells that are all rather similar except for the basal cell, which is modified into a holdfast. Filament cells may produce asexual zoospores mitotically; these have lour flagella, swim briefly, then settle and grow into new filaments. During sexual reproduction, some cells produce gametes, which can be identified because they have only two flagella, not four. Ulothrix is isogamous, and the gametes, winch all look like cells of Chlamydomonas, pair and fuse. The zygote germinates by meiosis, producing lour haploid zoospores, each of which swims for a period, then loses its flagella, attaches to a substrate, and grows into a new filament.
Spirogyra is an extremely common fresh-water filamentous green alga (Fig. 11b) found in streams and ponds throughout North America. Its cells have beautiful spiral band-shaped chloroplasts that wind around the cell just below the plasma membrane. Swimming gametes are not formed; instead filaments undergo conjugation. Each filament is haploid and is either a + or — mating type. If compatible filaments drift against each other, a conjugation tube forms between cells, and the — protoplasts migrate through the tube and fuse with the + protoplasts. Karyogamy follows, although as many as 30 days may pass before nuclei fuse. The cell becomes dormant, thick-walled, and resistant; it later germinates and grows into a new filament. Meiosis occurs immediately after karyogamy, so the spore is haploid.
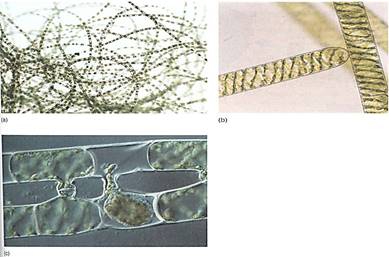
FIGURE 11: (a) Individuals of Ulothrix are uniseriate, unbranched filaments of haploid cells. When the spores or gametes are released, only empty cell walls remain. Sporogenous cells do not have a protective layer of sterile cells surrounding them, whereas the sporogenous cells of flowering plants are surrounded by either the anther wall or the nucellus (X 80). (E. R Degginger) (b) Cells of Spirogyra, showing the spiral, band-shaped chloroplasts. (Runk/Schoenberger from Grant Heilman) (c) Compatible filaments of Spirogyra have been brought together in culture and are undergoing conjugation. This type of plasmogamy is unusual in algae, being analogous to the plasmogamy of fungal hyphae. It superficially resembles conjugation in bacteria, but few details are similar. (M. 1. Walker/Science Source/Photo Researchers, Inc.).
Laminar Species. Ulva is slightly more complex than Ulothrix, but many of its stages are almost identical (Fig. 12). A quadriflagellate, haploid zoospore settles down and grows into a Ulothrix-like filament. The cells divide in two directions and thus form a sheet; then all cells divide once in a third direction so that the sheet becomes two layers thick, Ulva has a dibiontic life cycle with an alternation of isomorphic generations (Fig.13). During sexual reproduction, gametophyte cells produce biflagellate anisogametes, the smaller gametes being produced on one gametophyte and the larger ones on a different gametophyte. Having two different types of individuals in one generation is dioecy, just as in flowering plants. The zygote grows into a filament and then into a double-layered sheet just like the gametophyte.

FIGURE 12 : Individuals of sea lettuce, Ulva, are sheets two cells thick. They grow to almost any length or width but do not become thicker. If cut or torn into several pieces, each continues to grow. (William E. Ferguson).
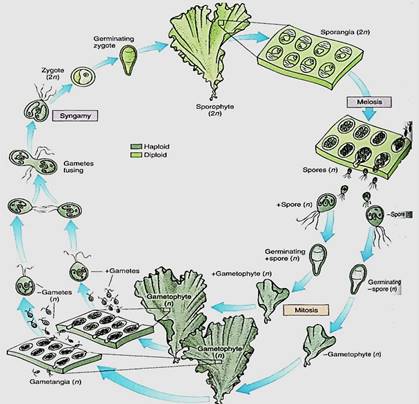
FIGURE 13:Life cycle of Ulva; details are given in text. Two multicellular, isomorphic generations occur. The types of reproductive structures and the size of the nuclei are the only reliable criteria £r distinguishing between sporophytes and gametophytes.
Coenocytic Species. A dibiontic life cycle with an alternation of heteromorphic generations is illustrated by Derbesia, the organism in which it was discovered (see Figs. 6b and 14). Before going into the details of the life cycle, think for a second about how organisms are studied. Often a biologist goes on a field trip, collects plants, insects, birds, or whatever, and preserves them by drying or fixing them in formaldehyde. It is difficult to maintain things in a living condition. Now imagine trying to do this 100 years ago when transportation was slow, laboratories did not have good artificial light or temperature control, and few pure chemicals were available for making culture solutions. Growing algae to study their life cycles was and is extremely difficult.
In 1938, P. Kornmann succeeded in obtaining zoospores from Derbesia marina, a branching, filamentous alga made up of giant coenocytic cells (Fig. 14). The zoospores were carefully maintained, but instead of growing into another Derbesia-like individual as expected, they grew into individuals of a totally different genus, Halicystis ovalis (Fig.14). A member of Halicystis looks nothing at all like Derbesia; instead it is composed of a single large, spherical coenocytic cell attached to the substrate by a small holdfast. Almost its entire volume is one giant vacuole, with only a thin layer of protoplasm next to the wall. When mature, Halicystis individuals, which are the gametophyte stage, produce either male or female anisogametes that undergo syngamy and establish the zygote. After germination, the zygote grows into a Derbesia sporophyte. To classify different life stages as different genera is obviously not correct, so the name Halicystis ovalis has been eliminated. Flowering plants also have an alternation of heteromorphic generations, but the gametophytes grow inside the sporophytes, so the question of which goes with which is easily solved.
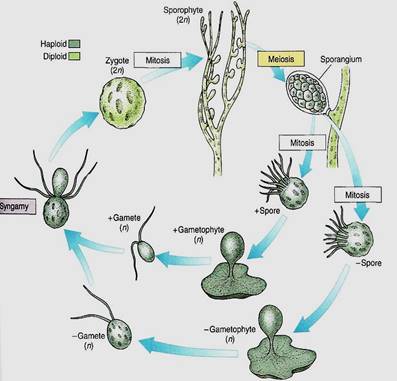
FIGURE 14:Derbesia has a dibiontic life cycle with an alternation of heteromorphic generations. The green algae are a large group (approximately 450 genera with 7500 species), and not all life cycles are known, so many more surprises of this type may be awaiting us. Only by growing all stages of a species can we study its life cycle.
Parenchymatous Species. Several groups of green algae undergo a true parenchymatous growth and may be related to the ancestors of true plants. One group, in class Chlorophyceae, divides by means of a phycoplast, which never occurs in plants, so this group is a remote possibility. The other group, in class Charophyceae, undergoes cell division by means of a phragmoplast, just as plant cells do. In Chlorophyceae, the flagellar root apparatus consists of four bands arranged in a cross; no true plant is known to have this type of basal body. In charophytes the flagellar root complex is similar to that of the motile cells of true plants: One major band of microtubules extends down into the cytoplasm from the basal body. Consequently, great attention is being given to charophytes.
An interesting example is Chara (Fig. 15), which has a stem-like body divided into nodes and internodes, with whorls of branches arising at internodes. The body is several cells thick, composed of true parenchyma tissue derived from cell division in all three planes and originating from an apical meristem that contains a prominent apical cell. Although these features seem to correspond to those of flowering plants, virtually all of the resemblance is spurious, because the earliest vascular land plants had no nodes, internodes, or branches . If Chara's ancestors were the ancestors of true plants, the only features that may be homologous rather than analogous are the parenchymatous body and growth by an apical meristem. Simple parenchymatous bodies also occur in members of Coleochaete, another group of charophytes that are being studied as possible close relatives of land plants.
Reproduction in Chara is significant. True plants (embryophytes) have multicellular reproductive structures with sterile cells, and on this basis Chara would have to be classified as a plant rather than an alga. Its sperm cells are produced in a truly multicellular gametangium whose outer cells are sterile; only the inner cells convert to sperm cells (Fig. 15b). At maturity, the outer cells separate slightly and the motile sperm cells swim away. The egg is formed as the terminal cell of a short filament three cells long, but the sub-terminal cell subdivides, and those cells grow upward and surround the egg (Fig. 15c). After fertilization, the sterile cells surrounding the fertilized egg deposit thickenings on their inner walls, those adjacent to the zygote. The resting structure thus consists not only of a thick-walled zygote but also protective sterile cells. When the resting cell germinates, it grows out as haploid filaments that soon establish an apical meristem and parenchymatic growth.
Multicellular gametangia with sterile cells have been used as the primary defining feature of embryophytes, true plants. Morphologists have long debated whether the gametangia of charophytes are ancestral to those of true plants or are merely a fascinating case of convergent evolution. Although they are similar, they are not identical, but Chara and Coleochaete are groups of living species, none of which is the direct ancestor of true plants which arose 420 million years ago. The question is whether they are members of an evolutionary line that has changed little since its ancestors gave rise to the ancestors of true plants, or whether they are only very distantly related, their parenchyma, apical growth, and multicellular gametangia all being cases of convergent evolution.
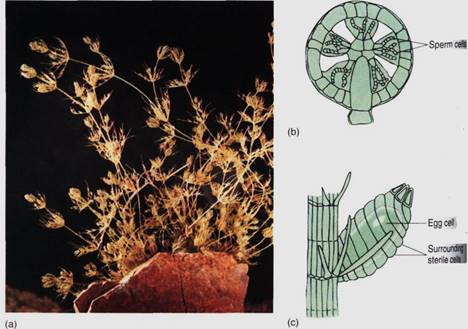
FIGURE 15: (a) An individual of Chara has a strong resemblance to certain aquatic angiosperms, but most of the similarity is analogous (resulting from convergent evolution), not homologous (resulting from descent from a common ancestor). (Runk/Schoenberger from Grant Heilman) (b) The microgametangium of Chara consists of an outer layer of sterile cells. Some inner cells mature into sperm cells, whereas others remain sterile and act as spacers. (c) The megagametangium is multicellular at maturity but not at initiation. The sterile cells around the egg are not sibling cells of the egg but rather are filaments that grow up from the cell below the egg. This is a multicellular gametangium, but it is filamentous, not parenchymatous.
 الاكثر قراءة في الطحالب
الاكثر قراءة في الطحالب
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة





















