























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Division Rhodophyta: Red Algae
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 17-11-2016
17-11-2016
 5129
5129




+
-
20
Division Rhodophyta: Red Algae
The red algae constitute a large group (about 400 genera and 3900 species) of especially distinct and fascinating algae (Figs. 1, and 2, and Table ). Numerous structural, biochemical, and reproductive features set them off from the other algae as well as from true plants. One of the most important biochemical distinctions of red algae is that, like cyanobacteria, they contain phycobilin accessory pigments that are aggregated into phycobilisomes . Their red color is due to the presence of phycoerythrin; however, they are often purple, brown, or black owing to the additional presence of phycocyanin, just as in cyanobacteria. Carotenoid accessory pigments are also present, as is chlorophyll a. The actual quantities of each type of pigment vary with depth. Those algae that grow in bright surface waters have an array of pigments suitable for relatively intense light, whereas algae in deeper waters have a different complex of pigments that is better suited both to the dimmer light and to the altered spectrum present as a result of the water's differential color absorption.

FIGURE 1:Many red algae have highly branched, filamentous bodies. Polysiphonia (X 20). (Runk/Schoenberger from Grant Heilman).
Excess photosynthate is stored as floridean starch, a branched polymer of glucose somewhat similar to glycogen; it occurs as granules in the cytoplasm, never in the chloroplast. Other reserves occur which contain unusual sugars such as floridoside and isofloridoside, indicating that the carbohydrate metabolism of red algae has characteristic features not shared by plants.
In addition to a thin layer of cellulose, the walls of red algae contain a thick layer of slimy mucilages composed of compounds called sulfated galactans. These are important commercially as thickening, suspending, or stabilizing agents in puddings, ice creams, cheeses, and salad dressings. The culture medium agar is also extracted from them. The complete wall—cellulose layer plus mucilage—is quite thick, often as thick as the protoplast is wide. Almost all red algae are multicellular, so a large fraction of the individual's volume consists of this apoplastic space. As a result, all cells, even the most internal ones, may have relatively direct contact with the surrounding water. In the largest family of rhodophytes, the Corallinaceae or coralline red algae, such large amounts of calcium carbonate are deposited in the walls that they become rocklike (Fig. 2). It was not until 1837 that they were recognized as algae rather than corals.
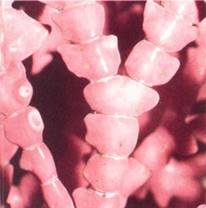
FIGURE 2 :Coralline red algae have walls so heavily impregnated with calcium carbonate that the body is hard and brittle. (Peter Arnold/Sea Studios, Inc.).

Walls of red algae lack plasmodesmata of the type that occurs in plants, but they do have distinctive pit connections . During cell division, the new wall is formed as a ring that grows inward from the original cell wall; the growth of the septum stops before it is complete, leaving a hole in the center. Vesicles then deposit a material that precipitates, forming a lens-shaped plug. Whether these are a means of intercellular transport or communication is not known, but when two separate filaments of cells come into contact, a "secondary pit connection" can form between them.
Red algae are usually multicellular; only a few unicellular species (Porphyridium, Rhodospora) have ever been discovered. Most red algae tend to be rather conspicuous, often Beautiful individuals and arc filamentous, membranous, or foliaceous; true parenchymatous bodies are reported to occur in only a few algae, such as Bangia and Porphyra. Typically red algae are smaller and less complex than brown algae. Like brown algae, the ancestors of the red algae must have diverged from the ancestors of green algae very early, certainly before multicellularity had evolved.
Despite the rather large size that the bodies of red algae can attain, little differentiation or specialization occurs among the cells. The greatest differentiation typically involves only cell size and pigmentation. Outer cells are smaller and more heavily pigmented; inner cells are larger and have fewer, smaller chloroplasts.
Most species grow attached by rhizoids to rocks, shells, algae, or sea grasses. Numerous red algae (more than 40 genera) are parasitic, usually on other red algae. The basal cell of these species penetrate into the host, forming secondary pit connections with host cells.
The life cycles of most red algae are poorly known, but the few that have been studied well are all extremely complex, almost all involving at least one multicellular stage but none having any motile cells; flagella and centrioles do not occur in any stage of any species. Many variations occur and there is no "typical" red alga life cycle. Figure 3 is a generalized life cycle. Gametophytes bear gametangia that produce nonmotile sperm cells called spermatia and egg-like cells called carpogonia. The carpogonia are large cells with a long tubular extension that basically acts as a receptor for drifting spermatia. When a spermatium contacts the extension, plasmogamy occurs and the nucleus migrates to to the carpogonium base, where karyogamy occurs. In any other group, this cell would be a zygote and would either grow or produce spores. But in many red algae, the fertilized carpogonium puts out another long filament that carries the diploid nucleus out of the carpogonium and deposits it into a totally different cell, an auxiliary cell, where mitosis begins and produces a mass of cells. This mass is a new generation, the carposporophyte, and it produces diploid carpospores by mitosis. The carposporophyte is a totally distinct generation from any other mentioned for any other algae or plants. Carpospores are released, float away and settle, and then germinate and grow into diploid plants called tetrasporophytes, equivalent to a regular sporophyte; these have sporangia in which the cells undergo meiosis, producing haploid tetraspores that grow into gametophytes.

FIGURE 3: Life cycle of a red alga; details are given in text. Note especially the lack of swimming cells and the presence of three generations, one haploid and two diploid.
 الاكثر قراءة في الطحالب
الاكثر قراءة في الطحالب
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















