























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Basic Types of Cells and Tissues
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 9-11-2016
9-11-2016
 5070
5070




+
-
20
Basic Types of Cells and Tissues
Despite the diversity of types of stems that have originated by natural selection, all share a basic, rather simple organization. The same is true for leaves and roots. Although we might suspect that numerous types of cells are present within a plant, actually the various kinds of plant cells are customarily grouped into three classes based on the nature of their walls: parenchyma, collenchyma, and sclerenchyma.
PARENCHYMA
Parenchyma cells have only primary walls that remain thin (Table ). Parenchyma tissue is a mass of parenchyma cells. This is the most common type of cell and tissue, constituting all soft parts of a plant. Parenchyma cells are active metabolically and usually remain alive once they mature. Numerous subtypes are specialized for particular tasks (Fig. 1).


FIGURE 1: (a) Parenchyma cells of geranium; their walls (green) are thin, and their vacuoles are large and full of watery contents that did not stain. Nuclei were present in all cells, but because these cells were so large and the section was cut so thin, most nuclei were cut away during the preparation of this slide. One nucleus is still present (X 160). (b) Chlorenchyma cells from a leaf of privet. Because these cells are small and the section is thicker than that in (a), most of these cells still have nuclei (red). The green structures close to the wall (blue) are chloroplasts. The large white areas are intracellular spaces where the cells have pulled away from each other. The spaces permit carbon dioxide to diffuse rapidly throughout the leaf (X 160). (c) Material taken from the center of a pine (Pinus) stem. The cells that have stained dark purple are filled with chemicals called tannins; these are bitter and deter insects from eating the tissue (X 50). (d) A resinal canal in a pine leaf. The white area is the central cavity where the resin is stored, and the cells that line the cavity are glandular parenchyma cells that synthesize and secrete the resin. The innermost cells have thin walls, which permit movement of resin from the cells to the cavity. The outer cells have thick walls, which provide strength; cells with thick walls are not parenchyma cells (X 160).
Chlorenchyma cells are parenchyma cells involved in photosynthesis; they have an abundance of chloroplasts, and the thinness of the wall is advantageous for allowing light and carbon dioxide to pass through to the chloroplasts. Other types of pigmented cells, as in flower petals and fruits, also must be parenchyma cells with thin walls that permit the pigments in the protoplasm to be seen.
Glandular cells that secrete nectar, fragrances, mucilage, resins, and oils are also parenchyma cells; they typically contain few chloroplasts but have elevated amounts of dictyosomes and endoplasmic reticulum. They must transport large quantities of sugar and minerals into themselves, transform them metabolically, then transport the product out.
Transfer cells are parenchyma cells that mediate the short-distance transport of material by means of a large, extensive plasma membrane capable of holding numerous molecular pumps. Unlike animal cells, plant cells cannot form folds or projections of their plasma membranes; instead, transfer cells increase their surface area by having extensive knobs, ridges, and other ingrowths on the inner surface of their walls (Fig. 2).
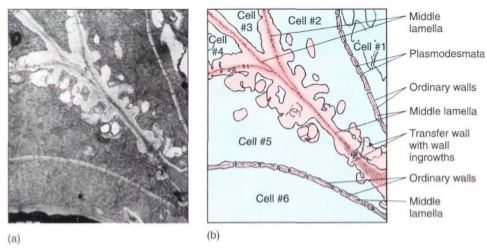
FIGURE 2: Transfer cells in the salt gland of Frankenia grandifolia. The wall ingrowths increase the surface area of the cell membrane, providing more room for salt-pumping proteins in the membrane (X 20,000).
ridges, and other ingrowths on the inner surface of their walls (Fig. 2). Because the plasma membrane follows the contour of all these, it is extensive and capable of large-scale molecular pumping.
Some parenchyma cells function by dying at maturity. Structures such as stamens and some fruits must open and release pollen or seeds; the opening may be formed by parenchyma cells that die and break down or are torn apart. Large spaces may be necessary inside the plant body; some of these are formed when the middle lamella decomposes and cells are released from their neighbors. In other cases, the space is formed by the degeneration of parenchyma cells. In a few species, such as milkweeds, as parenchyma cells die, their protoplasm is converted metabolically into mucilage or a milky latex.
Parenchyma tissue that conducts nutrients over long distances is phloem; it is discussed later in this chapter.
Parenchyma cells are relatively inexpensive to build because little glucose is expended in constructing the wall's cellulose and hemicellulose. Each molecule incorporated into a wall polymer cannot be used for other purposes such as the generation of ATP or the synthesis of proteins. Consequently, it is disadvantageous to use a cell with thick walls any time one with thin walls would be just as functional. Most leaves are soft, composed almost entirely of parenchyma, and are therefore not very expensive metabolically. After several weeks of photosynthesis they replace the sugar used in their construction, and all photosynthesis after that point is net gain for the plant.
COLLENCHYMA
Collenchyma cells have a primary wall that remains thin in some areas but becomes thickened in other areas, most often in the corners (Fig. 3). The nature of this wall is important in understanding why it exists and how it functions in the plant. Like clay, the wall of collenchyma exhibits plasticity, the ability to be deformed by pressure or tension and to retain the new shape even if the pressure or tension ceases. Collenchyma is present in elongating shoot tips that must be long and flexible, such as those of vining plants like grapes, as a layer just under the epidermis or as bands located next to vascular bundles, making the tips stronger and more resistant to breaking (Fig. 4). But the tips are still capable of elongating because collenchyma can be stretched. In species whose shoot tips are composed only of weak parenchyma, the tips are flexible and delicate and often can be damaged by wind; the elongating portion must be very short or it simply buckles under its own weight.
It is important to think about the method by which collenchyma provides support. If a vine or other collenchyma-rich tissue is cut off from its water supply, it wilts and droops; the collenchyma is unable to hold up the stem. Parenchyma cells are needed in the inner tissues for support. Collenchyma and turgid parenchyma work together like air pressure and a tire: The tire or inner tube is extremely strong but is useless for support without air pressure. Similarly, air pressure is useless unless it is confined by a container. In stems, the tendency for parenchyma to expand is counterbalanced by the resistance of the collenchyma, and the stem becomes rigid. Because the walls of collenchyma cells are thick, they require more glucose for their production. Collenchyma is usually produced only in shoot tips and young petioles, where the need for extra strength justifies the metabolic cost. Subterranean shoots and roots do not need collenchyma because soil provides support, but the aerial roots of epiphytes such as orchids and philodendrons have a thick layer of collenchyma.
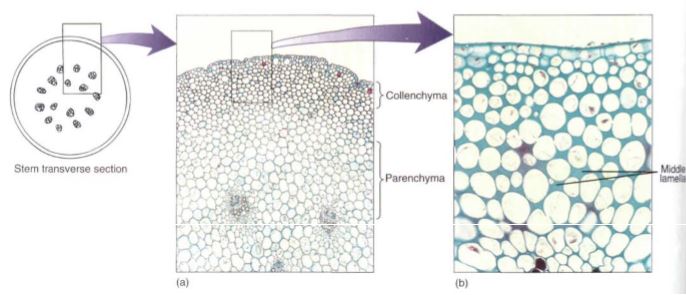
FIGURE 3: (a) Masses of collenchyma cells often occur in the outer parts of stems and leaf stalks; this is part of a Peperomia stem. The collenchyma forms a band about 8 to 12 cells thick. The inner part of the stem is mostly parenchyma (X 50). (b) Look closely between the cells and notice the fine dark lines—the original, thin primary walls and middle lamellas. In collenchyma cells, the primary wall gradually becomes thicker at the corners so the protoplast becomes rounded. No intercellular spaces are present (X 150).

FIGURE 4: The shoot tips of long vines need the plastic support of collenchyma while their stems are elongating.
SCLERENCHYMA
The third basic type of cell and tissue, sclerenchyma, has both a primary wall and a thick secondary wall that is almost always lignified (Fig. 5). These walls have the property of elasticity: They can be deformed, but they snap back to their original size and shape when the pressure or tension is released. Sclerenchyma cells develop mainly in mature organs that have stopped growing and have achieved their proper size and shape. Deforming forces such as wind, animals, or snow would probably be detrimental.
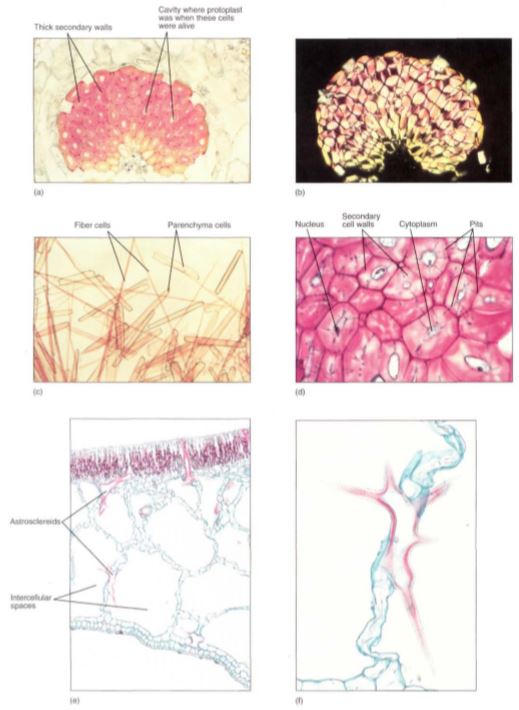
FIGURE 5.10 :(a) A mass of fiber cells in the leaf of Agave. These are large, heavy, tough leaves, and the fiber masses give the leaves strength. Notice that each cell consists mostly of thick secondary cell wall; the small white space in each is an area where the protoplast had been before it died (X 150). (b) This is the same mass of fibers as in (a) but viewed with polarized light. The thick secondary wall shines brightly because its cellulose molecules are packed in a tight, crystalline form, giving the wall extra strength (X 150). (c) A stem of bamboo was treated with a mixture of nitric acid and chromic acid to dissolve the middle lamellas and allow the cells to separate from each other. In this preparation you can see that the fibers are long and narrow. The shorter, wider cells are parenchyma (X 80). (d) These are sclereids; they are more or less cuboidal, definitely not long like fibers. These have remained alive at maturity, and nuclei and cytoplasm are visible in several. The blue-stained channels that cross the walls are pits with cytoplasm. The pits of each cell connect with those of the surrounding cells so that nutrients can be transferred from cell to cell, keeping them alive (X 150). (e) This portion of a leaf of water lily contains large, irregularly branched cells that have stained red. These are known as astrosclereids (star-shaped sclereids). The large white spaces are giant intercellular spaces; this is an aerenchyma type of parenchyma (X 40). They are shown at higher magnification in (f). (f) A star-shaped sclereid, showing only part of its long, arm-like extensions. The ends of most of the extensions were cut off when the material was cut to make this slide. Tiny cubic crystals are present in the wall (X 150).
If mature organs had collenchyma for support, they would be reshaped constantly by storms or animals, which of course would not be optimal. For example, while growing and elongating, a young leaf must be supported by collenchyma if it is to continue to grow. But once it has achieved its mature size and shape, some cells of the leaf can mature into sclerenchyma and provide elastic support that maintains the leafs shape. Unlike collenchyma, sclerenchyma supports the plant by its strength alone; if sclerenchyma-rich stems are allowed to wilt, they remain upright and do not droop.
Parenchyma and collenchyma cells can absorb water so powerfully that they swell and stretch the wall, thereby growing; sclerenchyma cell walls are strong enough to prevent the protoplast from expanding. The rigidity of sclerenchyma makes it unusable for growing shoot tips because it would prevent further shoot elongation.
Sclerenchyma cells are of two types—conducting sclerenchyma and mechanical sclerenchyma. The latter type is subdivided into long fibers (Fig. 5a to c) and short sclereids (Fig. 5d to f; Table 2), both of which have thick secondary walls. Because fibers are long, they are flexible and are most often found in areas where strength and elasticity are important.

The wood of most flowering plants contains abundant fibers, and their strength supports the tree while their elasticity allows the trunk and branches to sway in the wind without breaking (usually) or becoming permanently bent (Fig. 6). The fiber-rich bark is important not in holding up the tree but in resisting insects, fungi, and other pests.
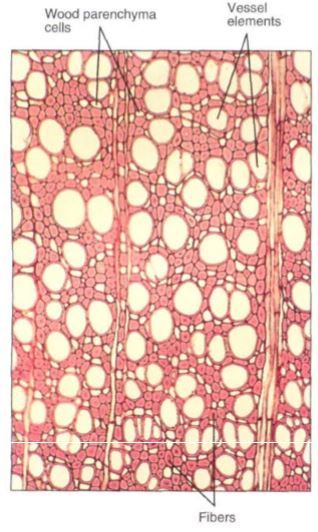
FIGURE 6: Wood is composed of several types of cells. The numerous small cells with thick walls and extremely narrow lumens are fibers. These give the wood strength and flexibility. The large round cells that appear to be empty are vessel elements, discussed later in this chapter; the small cells with thin walls and large lumens are wood parenchyma cells (X 60).
Sclereids are short and more or less isodiametric (cuboidal). Because sclereids have strong walls oriented in all three dimensions, sclerenchyma tissue composed of sclereids is brittle and inflexible. Masses of sclereids form hard, impenetrable surfaces such as the shells of walnuts and coconuts or the "pits" or "stones" of cherries and peaches. Flexibility there would be disadvantageous because the soft seed inside might be crushed even though the shell remained unbroken.
When strength or resistance is the only selective advantage of sclerenchyma, the protoplast usually dies once the secondary wall has been deposited. But in some species, certain sclerenchyma cells, especially fibers, remain alive at maturity and carry out an active metabolism (Fig. 7). These living sclerenchyma cells most often are involved in storing starch or crystals of calcium oxalate. Some have rather thin secondary walls, but in others the secondary walls are just as thick as those of fibers that die at maturity and provide only support.
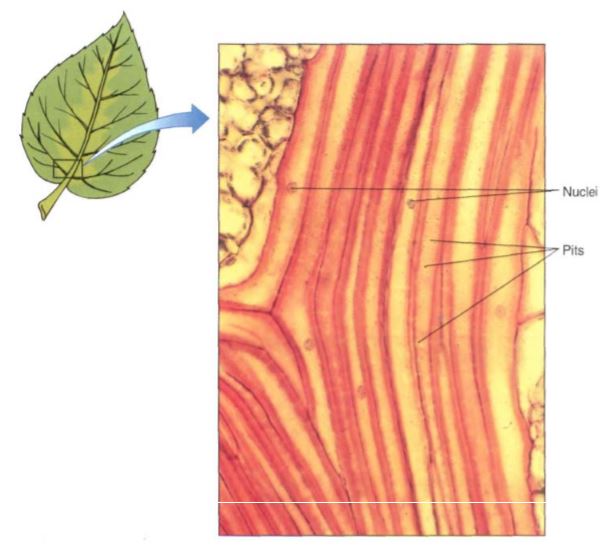
FIGURE 7: These fiber cells have nuclei, indicating that they are living cells. The secondary walls are thick, but not so thick as the walls in Figure 5.11. The small dots visible in the walls are pits; these are much narrower than the pits of Figure 5. 10d. Leaf of Smilax (X 150).
Like all cells, sclereids and fibers develop from cells produced by cell division; when newly formed, they are small and have only a primary wall—they are parenchyma cells. If the cell is to differentiate into a sclereid, it may expand only slightly, but if it is to develop into a fiber, it elongates greatly. When immature sclereids and fibers reach their final size, the cellulose-synthesizing rosettes of the plasma membrane begin to deposit the secondary wall. As this wall becomes thicker and is impregnated with lignin, it becomes waterproof, so nutrients can enter the cell only through plasmodesmata rather than everywhere, as with, parenchyma and collenchyma cells. It is important that the secondary wall not be deposited along the entire inner surface of the primary wall; small, plasmodesmata-rich areas must remain free of the secondary wall. At first these areas are low depressions in the developing secondary wall, but as wall deposition continues these areas become narrow pits in the secondary wall (see Figs. 5d). The pits of adjacent sclerenchyma cells must meet; if the pits of one cell met areas of secondary wall in the neighboring cells, no water or sugars could be transferred and the protoplasts would starve.
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















