























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Long Distance Transport : Xylem
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 2-11-2016
2-11-2016
 3835
3835




+
-
20
Long Distance Transport : Xylem
PROPERTIES OF WATER
The movement of water through xylem is based on a few simple properties of water and solutions. One property is that water molecules interact strongly with other water molecules, behaving as if weakly bound together; when frozen, the molecules become strongly bound to each other. Because of this, liquid water is said to be cohesive, and any force acting on one molecule acts on all neighboring ones as well.
Another property of water is that its molecules interact with many other substances- it is adhesive. Almost all substances in plants, except lipids, interact with water; water molecules adhere to these substances and cannot be easily removed from them. Cellulose molecules, enzymes, DNA, sugars, and so forth, have a shell of water molecules rather firmly attached to them. Occasionally a water molecule vibrates out of one of these shells and is replaced by another water molecule, but in general, adhesion makes these water molecules less free to move around than other water molecules.
Water also adheres firmly to soil particles. When soil is quite moist, roots can absorb the free liquid water between soil particles. But as the soil dries, the remaining water adheres firmly to the soil and cannot be absorbed easily, if at all (Fig. 1). Even though the soil may contain considerable water, it is unavailable to the plant. The same is true of seawater; the water molecules interact so strongly with the salt molecules that land plants cannot pull the water away. Another property of water is that it is heavy, and lifting it to the top of a tree requires a great deal of energy. If water were lighter, less energy would be involved.
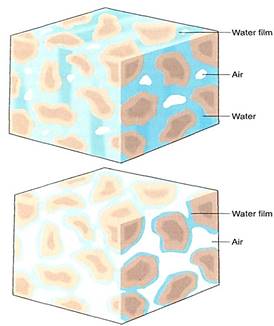
FIGURE 1:Wet soil contains water both as a film covering all surfaces of soil particles and as small mass held in the capillary spaces formed where two soil particles touch each other (a). The latter is held weakly and can be easy absorbed by roots. (b) Dry soil contains only the tightly bound films of water. This adhesion is measured by ym, which can be so negative that the soil's wafer potential is also very negative. This water cannot be absorbed by the roots of most plants.
WATER TRANSPORT THROUGH XYLEM
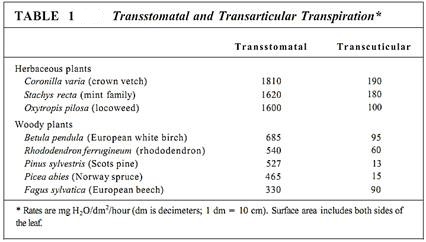
Water movement through xylem and plants as a whole is governed by the principles of water relations just described. The cohesion-tension hypothesis is the most widely accepted model of the process. When stomatal pores are open, they unavoidably allow water loss. The apoplastic space of spongy mesophyll and palisade parenchyma is filled with moisture-saturated air, so water molecules have a strong tendency to diffuse from intercellular spaces (the point of less negative water potential) to the atmosphere (the point of more negative water potential). This water loss is called transstomatal transpiration. The cuticle and waxes on the epidermal surfaces are fairly efficient isolation mechanisms, being so hydrophobic that very little water passes through them. However, some water is lost directly through the cuticle by transcuticular transpiration (Table 1).
Consider a leaf in early morning: Stomata are closed, air is cool, and relative humidity is high. The air may have cooled enough during the night to allow dew to form. Cells within the leaf are turgid and probably in equilibrium with each other, all having a water potential between 0.0 and — 1.0 MPa . As the sun rises, stomata open and begin losing water; the air warms and its relative humidity decreases. As transpiration causes epidermal cells and mesophyll cells near stomata to lose water, their water potentials become more negative, going out of equilibrium with surrounding cells. The disequilibrium does not become major because water diffuses into these cells from other cells and apoplastic spaces deeper within the leaf. But this water movement out of the deeper mesophyll cells causes their water potentials to become more negative, away from equilibrium with even deeper cells (Fig. 2).
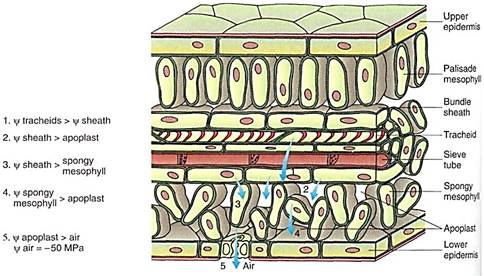
FIGURE 2:As water moves out of the leaf into the air, the tissues dry and a water potential gradient becomes established. Water flows from the xylem, where water potential is least negative, toward air, where water potential is most negative.
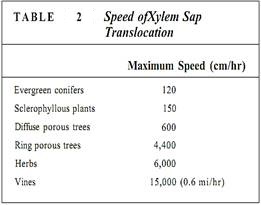
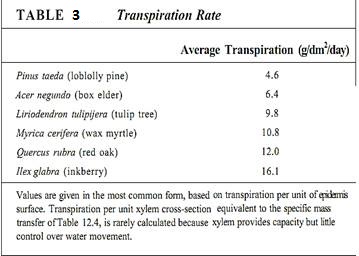
Finally, this gradient of water potentials reaches a tracheid or vessel member. As water molecules move out of tracheary elements into mesophyll parenchyma cells, the water potential within the xylem water column becomes more negative. The loss of water from tracheary elements does not really affect the xylem osmotic potential because solutes are very dilute to begin with. Here water's cohesive properties are more important: As a water molecule leaves the xylem, it does not leave a hole behind but instead drags other water molecules along with it. All water molecules of the plant are hydrogen bonded together, but the water molecules in the xylem can move upward most easily. That water is purest, is not bound to proteins and cellulose, is not locked into hydration shells around solutes, and so on. As water molecules diffuse out of xylem in the leaves, cohesive forces pull water upward through the xylem, all the way from the roots (Tables 2 and 3). Imagine the water being frozen solid; if the top molecules are pulled upward, the entire mass of ice is lifted.
Water is heavy, and water molecules in the uppermost tracheary elements must lift the weight of the entire water column. There is tension on these molecules, and consequently the pressure potential is a negative number; as water moves into the leaf mesophyll, the xylem water potential becomes more negative owing to an increasingly negative pressure potential. In vertical stems, water must move directly upward in the xylem, and the water's weight is a significant consideration, fn the examples discussed earlier, water could move laterally between a cell and a beaker, so no lifting was involved. In vertical xylem, the weight of water counteracts its tendency to rise into areas of more negative water potential.
Consequently, if leaf xylem water potential is only slightly more negative than root xylem water potential, the water does not move. For every 10 meters of height, leaf water potential must be at least 0.1 MPa more negative than root water potential (Fig. 3). In trees such as elms and sycamores that are typically more than 30 meters tall, leaf water potential must be at least 0.3 MPa more negative than root water potential simply to overcome the weight of water. This is accomplished automatically: When stomata open in the morning, leaf cells lose water and their water potentials become more negative. However, water does not begin moving upward in the xylem until drying causes the water potential of leaf cells to become sufficiently more negative. Stolons, rhizomes, and horizontal vines have no such problem; long distance transport is horizontal and no lifting is involved, so gravity is not a factor.
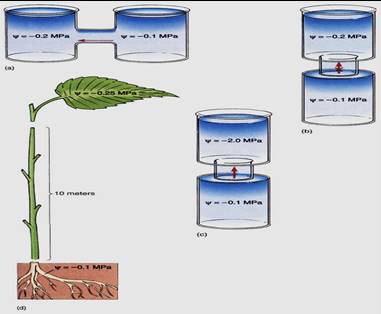
FIGURE 3: (a) If water can move laterally between two solutions, no lifting is involved. The slightest difference in water potential results in water movement. (b) If water must move upward against gravity, work must be done. A slight difference in water potential may not be enough to cause water movement; only a large difference will (c). (d) To overcome gravity and friction, the water potential of plant tissues receiving water must be at least 0.2 MPa more negative than that of roots for every 10 meters of height separating them. In the case illustrated here, with a difference of 0.15 MPa, water would not move up the stem. The water potential of the leaf would have to become 0.5 bars more negative.
A few plants grow as pendant epiphytes, their stems dangling down from the branches of the host plant. Their stems and leaves are lower than their roots, and gravity assists water movement (Fig. 4).

FIGURE 4:Because the transpiration surfaces of this epiphytic cactus (Rhipsalis) are lower than its roots, water flows downward from the roots and gravity actually assists xylem conduction rather than hindering it, as in most plants.
Water is extremely adhesive, and its molecules interact strongly with the polymers of the cell walls of tracheids and vessel elements. Water molecules adjacent to the walls tend to remain fixed to the walls and also tend to prevent neighboring water molecules from being drawn upward by transpiration/cohesion. This results in a layer of relatively immobile water that does not move easily. In narrow tracheids and vessel elements, this immobile water is a significant fraction of the water column. The resulting friction hinders water’s movement and contributes to the tendency of water to not move upward even when leaves have a more negative water potential than roots. As a rough approximation, to overcome friction, leaf water potential must be at least 0.1 MPa more negative than root water potential for every 10 meters of height. Therefore, considering both friction and gravity, a difference of 0.2 MPa is needed for every 10 meters of height. In plants with numerous wide vessels, friction is less, and less than 0.2 MPa is needed, but in plants with narrow tracheary elements, even more than 0.2 MPa is necessary. Also, in plants that have only tracheids, the water molecules must be pulled through pit membranes when entering and leaving each tracheid, further contributing to friction.
Returning to the plant in our example, transpiration causes leaf cells to lose water and their water potentials become more negative; water moves into them from tracheary elements and tension builds on water molecules in the xylem. When the water potential in the uppermost tracheary elements has become sufficiently more negative than that in the lower elements, friction and gravity are overcome and water moves upward. This causes the lowermost xylem cells to pull water inward from the root cortex. The cortex water potential drops lower than that of the root epidermis and root hairs, and water flows from them to the cortex. In the final step, the water potential of the root epidermis and root hairs. becomes more negative than that of the soil, and water moves automatically into the root from the soil.
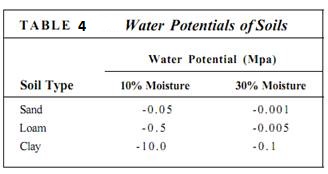
Long distance water transport occurs in this manner as long as the soil is sufficiently moist. A sandy soil that has 30% moisture has a water potential of about — 0.001 ME% almost equal to that of pure water (Table 4). Water is held in soils by cohesion and adhesion as wedges and droplets between soil particles, and in sandy soils, root hairs can easily draw water from moist soils. Gravity and evaporation also pull water away from the droplets in sandy soils, so these soils dry quickly after a rain. As the soil dries, the most mobile molecules are removed and those tightly bound to soil particles remain . In a dry soil, not only is less water present, but it is also relatively immobile. Clay soils are composed of thin flakes with a high surface-to-volume ratio. When wet, they hold a great deal of water, but it is firmly bound as a hydration layer. No root can pull water away from clay that is even slightly dry. Loam soils consist of sand, silt, and clay and have a diversity of pore sizes. During a rainfall, loam soils absorb large amounts of water, then hold it for weeks after the rain has stopped. The diversity of pore sizes allows root hairs to absorb water but prevents gravity from pulling the water so deep into the soil that roots cannot reach it.
Many roots remain healthy with their water potentials as low as —2.0 MPa; they still absorb water from soils that are quite dry (Fig. 5). But if the plant is 10 meters tall, the leaves would have to have a water potential at least slightly more negative than -2.2 MPa to overcome friction and gravity. This is also possible in some species, but typically the leaves would either be dormant or preparing for abscission.

FIGURE 5: (a) Mary desert shrubs withstand severe desiccation; even though their water potentials become extremely negative, the cells survive, although they may become inactive. (b) Plants such as this mash plant (Nymphoides peltata) do not tolerate water stress at all; if their water potential falls very far below —0.2 MPa, they die quickly.
When both soil and air are dry, plants are greatly stressed. Even if stomata close, transpiration continues, at least through the cuticle. Leaf water potentials become more negative, but water cannot move upward easily because the soil is so dry. Tension on the water columns increases and at some point cohesion is overcome: Hydrogen bonding is broken over a large region and the water column breaks. This breaking is called cavitation, and the water column acts just like a broken cable. Molecules above the cavitation point are drawn rapidly upward because they are now free of the weight of the water below them; those below the cavitation point rush downward because nothing supports their weight. Between the two portions is space called an embolism, which expands until its surface encounters a solid barrier such as a pit membrane. The water/embolism surface cannot pass through pit membranes, but it can pass through perforations because they are open holes (Fig. 6). When an embolism forms in a tracheid, only that tracheid loses its water, but when an embolism forms in a vessel element, it may expand through perforation after perforation until the entire vessel has been emptied.
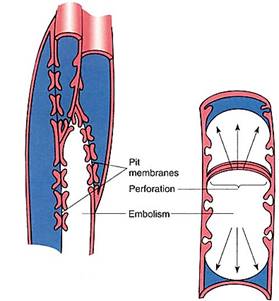
FIGURE 6:Severe tension can overcome the cohesion of water molecules and cause an embolism to form and expand rapidly. The embolism can pass through holes such as perforations but is stopped by pit membranes. If an embolism occurs in a tracheid, it cannot spread beyond the tracheid; but if a vessel element cavitates, the embolism spreads throughout the entire vessel.
Cavitation usually means that that tracheid or vessel can never conduct water again. The cohesive bonding that permits leaf transpiration to draw water upward has been disrupted. Under unusual conditions, embolisms are occasionally "healed." If all the surrounding cells are full of water and if the night is so cool and humid that transpiration stops, enough water may seep into the embolism to fill it and re-establish a continuous water column. More typically, any water that seeps in simply flows down the side of the tracheary element but is not able to fill it.
Because cavitation destroys the usefulness of an entire vessel or tracheid and because a plant invests considerable energy and reduced carbon in making tracheary elements, features that minimize cavitation are selectively advantageous. Adhesion between water and the cell wall is just such a feature, giving the water column extra strength so that it does not cavitate easily. This is most effective in narrow elements, where the reinforcement affects all water molecules, even those in the center. In wide elements, the central molecules are freely mobile and cavitate almost as easily as pure water.
This wall-induced reinforcement of water columns is believed to be the feature that allows plants to reach the heights they do. Eucalyptus trees in Australia are the tallest parts known; they grow to 100 meters in height, and water is pulled upward the entire distance through their tracheary elements. This cannot be duplicated with glass or metal capillary tubes; the water columns are too fragile to support their own weight without the reinforcing that cell walls provide. One hundred meters appears to be the limit for xylem; even with reinforcement, the cohesive forces at the top of the water column cannot support the weight of 100 meters of water hanging from them.
Some remarkably exquisite types of wood have evolved that are elegantly adapted for the various conditions in which plants live. In the moist tropics, water is always abundant, the soil is never dry, and water always moves easily; reinforcement is not necessary and the wood is full of wide vessels (Fig. 7). In drier temperate areas, especially rocky slopes, water is frequently scarce and water stress common. It is selectively advantageous for plants in such an environment to have narrow vessels or even wood with tracheids only, as conifers have. In temperate areas with good rainfall, the plants usually have a moist spring and produce early wood (also called spring wood) with large vessels; the summer is drier and they then produce late wood (summer wood) with narrower vessels or only tracheids. During the summer, the wide vessels of the early wood cavitate, and conduction occurs primarily or entirely in the late wood (Fig. 8).
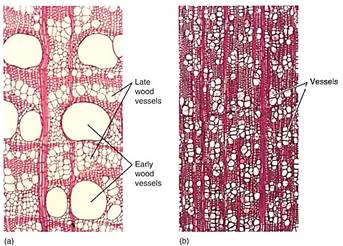
FIGURE 7: (a) Wood from a tropical tree. There are many broad vessels in the early wood, each of which can conduct water rapidly from the moist soil. Late wood has narrower vessels (X 100). (b) This wood is from a tree of temperate climates; the vessels are narrow and abundant. No single vessel can conduct very much water, but if one cavitates, only a small fraction of the conducting capacity of the wood is lost (X 100).
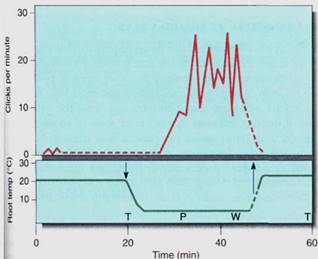
FIGURE 8:Cavitation of vessels and tracheids causes audible clicks that can be heard by sensitive microphones and counted. This graph shows the induction of cavitation in castor bean xylem; plants were grown in water solutions that permitted water uptake to match transpiration loss—there was little tension on xylem water columns and no cavitations occurred. Then the roots were cooled (arrow at left) to 5°C to inhibit root absorption of water, but shoots were kept warm to encourage transpiration. Water columns were stretched and immediately began breaking. After 20 minutes, embolisms were forming at almost 30 per minute and the plants were wilting. At 45 minutes the root solution was warmed to permit water absorption, and cavitation quickly stopped.
Eventually all vessels and tracheids cavitate. Dry conditions in summer cause many cavitations, as do freezing in winter, vibration in wind, and damage by burrowing insects. After tracheary elements cavitate, surrounding parenchyma cells may block them off by secreting gums and resins. If the tracheary element has wide pits, the adjacent parenchyma cells can soften the wall and swell into the vessel, blocking it with a mass of protoplasm called a tylosis. As more tracheary elements cavitate in a region of wood, the adjacent parenchyma synthesizes phenolic compounds that are resistant to attack by bacteria and fungi. These chemicals are often dark and aromatic, causing the wood to become brown and fragrant. The parenchyma cells then also die, and the region becomes part of the heartwood .
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















