























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Hormones as Signals of Environmental Factors
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 30-10-2016
30-10-2016
 4583
4583




+
-
20
Hormones as Signals of Environmental Factors
LEAF ABSCISSION
Whereas normal growth of shoots and roots results in large flows of auxin and cytokinin, respectively, environmental factors also influence hormone concentrations. Hormones communicate to various parts of a plant the information that a particular part has encountered an environmental change. Export of ABA by wilted leaves has been mentioned, and another example involves abscission of leaves and fruits. A young leaf produces large amounts of auxin, but production falls to a low but steady level in a mature leaf. As long as auxin flows out through the petiole, activity in the abscission zone is inhibited (Fig. 1). If the leaf is damaged by animal feeding or water stress, auxin production drops to such a low level that its flow through the petiole does not keep the abscission zone quiescent. Perception and transduction in this case may be simply that insect or wilt damage makes it impossible for the impaired cells to produce enough auxin to inhibit the abscission. Old age of the leaf may also result in the lack of sufficient auxin, but evidence suggests that autumn conditions stimulate production of ethylene, which then suppresses auxin production and transport in time for abscission before winter.
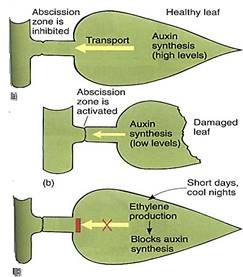
FIGURE 1: (a) A healthy leaf produces and transports enough auxin to suppress activity in the abscission zone. (b) A damaged leaf produces less auxin, insufficient to prevent abscission. (c) Autumn stimuli may cause the production of ethylene, which then suppresses auxin synthesis and transport. An alternative hypothesis postulates that autumn conditions directly suppress auxin production and ethylene is not necessary.
Fruits are prevented from abscising prematurely by the presence and export of sufficient amounts of auxin through the pedicel. Fruit ripening is under the control of both auxin and ethylene, at least in edible, fleshy fruits. Initial transformation of the ovary wall into a fruit is a response to auxin synthesized in developing embryos and transported to the ovary wall (Fig.2). Auxin stimulates many changes, including cell enlargement and differentiation; there is usually surprisingly little cell division during the formation of fruits, even large ones. One effect of auxin is release of ethylene by the developing fruits, which leads to other aspects of ripening in both climacteric and nonclimacteric fruits. At maturity the high concentration of ethylene stimulates the pedicel abscission zone, overriding the presence of auxin.

FIGURE 2 :Auxin's effects on fruit ripening are often studied in the strawberry because the tiny true fruits (achenes) are located on the outside of the large red false fruit. The achenes are sources of auxin necessary for development of the false fruit. If they are removed, no development occurs unless auxin is applied experimentally.
TROPISMS
Bending of plant parts toward or away from stimuli requires differential growth; that is, one side of the responding organ grows more rapidly than the other. In phototropism, the mechanism responsible is asymmetrical distribution of auxin. In positive phototropism of oat seedlings, the tip of the outermost, protective leaf, the coleoptile, is the site of perception; if it is covered with an opaque hood, light direction is not detected. The site of response, where differential growth causes bending, is about 5 mm below the coleoptile tip (Fig. 3). It has not been possible to determine conclusively which pigment is the photoreceptor for phototropism, but the result of unilateral illumination is a redistribution of auxin to the darker side of the coleoptile apex (Fig. 4). Careful measurements have shown that auxin synthesis is not affected; the gradient is not established by destruction of auxin on the lighted side or extra synthesis on the darker side. Once the differential distribution of auxin is established, the darkened side of a coleoptile, or the stem in other plants, receives extra auxin, so it grows more rapidly and the stem bends toward the light. When the stem points directly at the light, neither side is brighter nor darker, so differential auxin transport stops and the stem grows straight ahead.

FIGURE 3: (a) When illuminated from directly above, oat seedlings grow upward. (b) When a young oat seedling is exposed to light from one side, its outermost sheathing leaf, the coleoptile, bends and grows toward the light If the coleoptile apex is covered (c) or cut away (d), no response occurs to unilateral illumination, so the tip is the site of perception. If the site of response is covered, bending occurs, so the site of response is not involved in perception at all. (e) In dark conditions or with overhead lighting, auxin is transported symmetrically down the coleoptile, causing equal amounts of growth everywhere. With unilateral illumination (f), auxin is redistributed, with the darker side transporting more auxin than the lighted side, so the darker side grows faster, resulting in curvature. (g) and (h) Auxin can be collected in small blocks of agar or other absorptive material and then placed asymmetrically on a decapitated coleoptile; the side receiving auxin grows, but the other side does not.
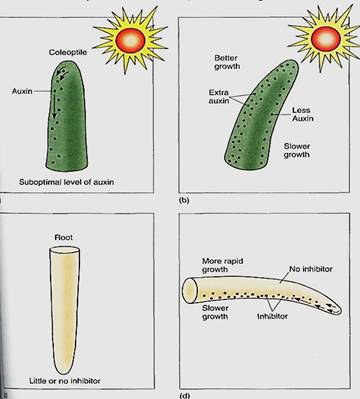
FIGURE 4: (a and b) Shoots may contain slightly too little auxin for fastest growth. Thus, redistribution in a coleoptile apex due to unilateral light causes the side with more auxin to have a level that is more nearly optimal, whereas the other side, which receives less, has even poorer growth than before. (c and d) In root positive gravitropism, the horizontal position of a root cap causes statoliths to fall to the side of statocytes, causing a downward redistribution of growth inhibitor. The side that receives inhibitor becomes inhibited, whereas the side that receives none has normal growth.
In positive gravitropism in roots, the root cap acts as the organ of perception .Once the lower side of the root is detected, a growth inhibitor is transported to that lower side. Unfortunately, we know almost nothing about how the position of the statoliths is transduced, nor do we know the identity of the growth inhibitor. Much circumstantial evidence indicates that it may be abscisic acid, but claims have been made for a different compound that has not yet been chemically identified.de of the root cap and then into the root, where it slows growth on the lower side.
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















