























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Activation and Inhibition of Shoots by Auxin
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 30-10-2016
30-10-2016
 3091
3091




+
-
20
Activation and Inhibition of Shoots by Auxin
Auxin is often described as a growth hormone, whereas abscisic acid is considered an inhibitor; unfortunately, such characterizations are confusing. Hormones simply carry information about the status of a particular region, nothing more; whether the elicited response is inhibition or activation depends on the site of response. An example of the complexity is provided by shoot tips.
As shoot apical meristems grow and initiate the new cells of shoots and leaf primordia, they also produce the auxin, IAA. Young leaves are also a rich source of this hormone. No external signal must be perceived to initiate auxin production; instead, this is a means of integrating the plant during ordinary growth. Large quantities of auxin indicate to cells that shoots are elongating and producing new leaves. Although neither signal perception nor transduction occurs, transport takes place. In stems auxin undergoes basipetal, polar transport through the cortex, perhaps by means of molecular pumps in plasma membranes. This downward flow of auxin surrounds all stem cells, and at least three cell types are set to respond to it, each response unique to the particular cell type.
CELL ELONGATION
In cells of the young internodes just below the apical meristem, auxin triggers cell elongation. When IAA contacts these responsive cells, which are prepared for growth, the crib begin to transport protons actively out across the plasma membrane (Fig. 1). This has the effect of acidifying the cell wall. The protons break some of the chemical bonds that hold one cellulose microfibril to another and activate enzymes that weaken other bonds so that the wall becomes weaker. If the protoplast is turgid and pressing against the wait it exerts enough pressure to stretch the weakened wall and growth results. Immature cells neither excrete protons nor grow if auxin is lacking. At lower internodes, fully grown. mature cells apparently lack the proper auxin receptors, because auxin does not cause them to extrude protons or grow.
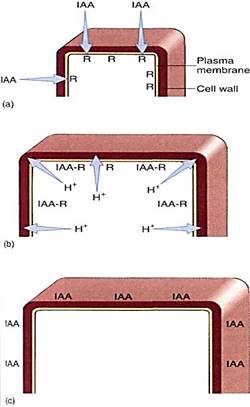
FIGURE 1 :Cells in the subapical region have auxin receptors. If the apex is growing and producing auxin, IAA is present to bind to the receptors (a). After stimulation by auxin binding, the plasma membrane pumps protons from the cytoplasm into the wall (b), weakening it and allowing turgor pressure to stretch it. (c) Cell elongation stops once the maximum cell size is reached, and adding more auxin does not cause any more elongation; perhaps the receptors have been removed from the membrane.
APICAL DOMINANCE
The second site of response to apically produced auxin is the buds located in leaf axils; their response is not cell elongation but rather inhibition of growth. Apically produced auxin induces dormancy in these axillary buds, the result being that each shoot tip has only one active apical meristem, a phenomenon called apical dominance (Fig. 2). This is a threshold response: As the terminal shoot apical meristem grows away, the concentration of auxin around an axillary bud gradually decreases until at some point it drops below the threshold. Inhibition cannot be maintained, and the axillary bud becomes active and grows out as a branch or flower. As the axillary bud grows, it produces auxin but does not inhibit itself, although it does inhibit all its own newly formed axillary buds.


FIGURE 2 :(a) Many cacti show almost 100% apical dominance; as long as the apex is undamaged, all axillary buds (spine clusters) remain dormant. Here, one bud has become active but is now producing enough auxin to keep its own buds dormant and suppress nearby buds on the parent stem. (b) The concentration of auxin is greatest at the shoot apex and less at lower levels; at sites where it drops below the threshold level, it can no longer suppress axillary buds, and one or several become active. Other factors must be involved, because it is not always the lowest bud that becomes active first, even though it should be the first to encounter sufficiently low auxin concentrations.
DIFFERENTIATION OF VASCULAR TISSUES
The third site of response to auxin produced in shoot tips is the vascular cambium; the response is cell division and morphogenesis. In springtime, as air temperatures rise and buds become active, their auxin moves basipetally, activating the dormant vascular cambium. Auxin not only stimulates cambial cells to begin mitosis and cytokinesis but also causes new daughter cells to differentiate into xylem cells. If an apical meristem is destroyed, by insects or a late frost for example, basipetal flow of auxin stops, vascular differentiation is interrupted, internode elongation ceases, and apical dominance is broken. Some axillary bud, now free of apical dominance, becomes active and re-establishes a flow of auxin that maintains the vascular cambium and any other cells that depend on it. Three separate target tissues give three distinct responses, not because there are three separate chemical messengers—there is only auxin—but because part of their previous differentiation was preparation to respond to auxin in a particular way. It is important to realize that the auxin carries no information except that the shoot apex is healthy and active. Each target site must have receptor molecules that interact with IAA and therefore detect the presence or absence of auxin. It is not known whether all three targets have the same receptors or if each has a unique type, nor is it known how the interaction of IAA with the receptor triggers the response.
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















