























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

The Structure of Ecosystems
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 20-10-2016
20-10-2016
 2980
2980




+
-
20
The Structure of Ecosystems
Many concepts can be considered in the structure of an ecosystem. Different botanists think about ecosystems in ways that are influenced by their own interests. Four of the most commonly mentioned structures are the physiognomic structure, temporal structure, species diversity, and trophic levels.
PHYSIOGNOMIC STRUCTURE
The physical size and shape of the organisms and their distribution in relation to each other and to the physical environment constitute the physiognomic structure. Trees, shrubs, and herbs are the three most useful categories, but in addition, a system of life forms was defined by C. Raunkiaer in 1934 (Table 26.3). The criterion for classification was the means by which the plant survives stressful seasons, such as by placing buds below ground (geophytes: bulbs, rhizomes) or winterizing aerial buds (phanerophytes: trees, vines). Regions of the world that have similar climatic conditions have similar physiognomic structures unless the soil is particularly unusual (Fig. 1).
An almost infinite number of combinations of life forms, vertical structures, and characteristics such as abundance of broadleaf plants, conifers, and sclerophyllous plants are possible (Fig. 2). An almost infinite number of types of ecosystems might be expected, but actually only a few basic types exist. Although differences exist between various types of forests or grasslands or marshes, each category is easily recognized as a common type of ecosystem.
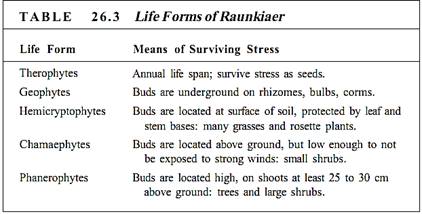

FIGURE 1: (a) This desert scrub vegetation with small trees, large bushes, and saguaro cactus is characteristic of much of the southwestern United States and northern Mexico, where summers are hot with occasional rainstorms and winters are cool and moist but freezes are not severe. (John Gerlach/Earth Scenes) (b) The northern part of the central valley in Chile, just north of Santiago, has climatic conditions similar to those in the southwestern United States, and the vegetation of the two regions resemble each other in habit (life forms), distribution, and other features. Only by looking carefully does one notice that the species are very different.
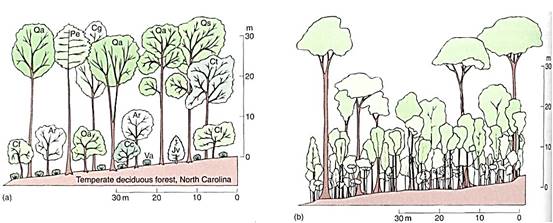
FIGURE 2: The presence of trees, shrubs, and herbs (phanerophytes, chamaephytes, and therophytes) gives an ecosystem vertical structure. The trees form the canopy; the shrubs, short trees, and saplings of tall trees form a middle level understory or subcanopy; the herbs and seedlings are the ground-level plants. Vines can occur in all three levels. (a) A temperate forest. Ar, Acer rubrum (red maple); Cc, Cercis canadensis (redbud); Cf, Cornus florida (dogwood); Cg, Carya glabra (pignut hickory); Ct, Carya tomentosa (mockernut hickory); Jv, Juniperus virginiana (red cedar); Oa, Oxydendrum arboreum (sourwood); Pe, Pinus echinata (shortleaf pine); Qa, Quercus alba (white oak); Qs, Quercus stellata (post oak); Va, Viburnum affine (arrow-wood). (b) A tropical rain forest has a much more complex structure. For more information about the research represented by these figures, see Plants, Man, and the Ecosystem by W. D. Billings and Tropical Rainforests of the Far East by T. C. Whitmore.
TEMPORAL STRUCTURE
The changes that an ecosystem undergoes with time constitute its temporal structure; the time span can be as short as a day or can encompass seasons or decades. For animals, a daily cycle can be especially obvious, with some animals active at night (nocturnal) and others during the day (diurnal). Many plants also have daily rhythms of flower opening and closing.
Plants change dramatically with the season, as do the other organisms. Spring is typically a time of renewed activity, the production of flowers and new leaves. This is not simultaneous for all species; often the understory plants become active earliest, benefitting from the open canopy. Wind-pollinated flowers are usually produced before leaves expand and block the wind. Leafing out and flowering must be coordinated not only with the end of low-temperature stress conditions but also with the habits of pollinators and herbivores. It does a plant no good to produce flowers when temperatures are still too low for its pollinators. Through late spring and early summer, various species flower at distinct times, controlled by plant maturity, photoperiod, or adequate rainfall. In most ecosystems there is no time during the summer when nothing is in bloom. The continual presence of some species in flower provides nectar and pollen for insects throughout the season.
Species that bloom early in spring probably form fruits and mature seeds during summer; species that flower later release their seeds in late summer or autumn. Once seeds are dispersed, some remain dormant until the following spring. Others germinate and grow into a low rosette that survives the winter, even growing slightly on warm winter days. When spring arrives, the seedling is already well-rooted and can begin to grow quickly while the seeds of competitors are just starting to germinate (Fig. 3).
Late summer and autumn bring changes that depend on the ecosystem; in the northern United States, herbs die while shrubs and trees develop resting buds. The entire plant enters light dormancy; then the first cool days initiate deep dormancy. Leaves and fruits are abscised, removing the last sources of food for most animals. In the southern and southwestern United States, cooler autumn weather is often more welcome than the first warm days of spring because the summer is so much more severe than the winter. Most gardening is done in autumn rather than spring, and fall wildflowers are abundant and dramatic. The growing season extends at least to December for shrubs and many herbs, and the small rosette plants may never become truly dormant.
In tropical ecosystems, winter and summer do not exist, but an alternation of dry and wet seasons governs ecosystem change. Coastal marsh, wetland, and reef ecosystems may be strongly affected by seasonality; rainy seasons dilute the salt water, whereas in dry seasons rivers deliver less fresh water and mineral-rich silt.
Over long periods—many years—most ecosystems undergo gradual, often dramatic changes.
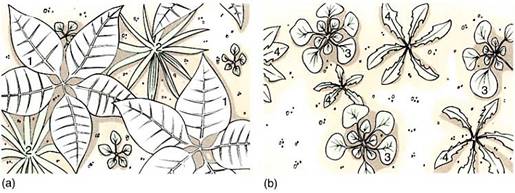
FIGURE 3: The temporal patchiness of an ecosystem is important; a large perennial plant occupies a site year after year, so the site is unavailable to seeds. But a site is occupied only temporarily by short-lived plants and becomes available for seeds when the plant dies. (a) Species 1 and 2 are winter rosettes that occupy most of the site through winter and early spring. (b) After they flower, fruit, and die, their sites are vacated and species 3 and 4, which germinate in later spring, find many sites available.
SPECIES COMPOSITION
Species composition refers to the number and diversity of species that coexist in an ecosystem, and it depends on whether the climate is mild or stressful, the soil is rich or poor, and the species' tolerance ranges are broad or narrow (Figs. 4 and 5). Stressful climates with poor soils support a low number of species, because so few species are adapted to such conditions (Fig. 6). On the other hand, mild climates and rich soils support an abundance of species because most plants have tolerance ranges that include such climatic and soil conditions (Fig. 7). Competition is intense, but apparently natural selection has resulted in habitat partitioning, with each species occupying a narrow portion of the various resource gradients (Fig.8). The presence of a large number of species actually creates more niches that can be filled by new species; the presence of trees makes it possible for epiphytes and parasites to occur in the ecosystem.
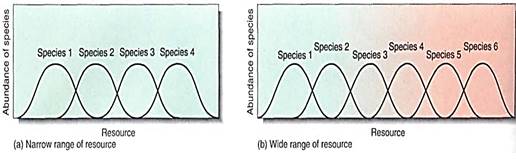
FIGURE 4: In the ecosystem represented by (a), the range of the resource available is narrow compared to the of the ecosystem in (b); in general, it is able to support fewer species. For example, the resource may be water, with the right ecosystem having a variety of areas that range from dry to moist to lakes or streams, whereas the left ecosystem is just marshy or just desert.
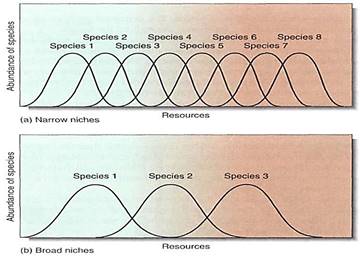
FIGURE 5: Two ecosystems with similar ranges of resources can differ in the number of species they contain. One may be occupied by a large number of very specialized species, each adapted to only a narrow range of the resource, whereas the other may be occupied by a few species of generalists that grow well under a variety of conditions and exclude most competitors.
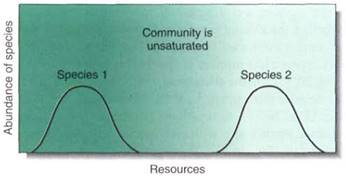
FIGURE 6: Adapting to certain harsh conditions is difficult, and only a few species may have succeeded. Consequently, harsh habitats may have many unused resources.

FIGURE 7: This climate is no stressful, the soil is rich, and no toxins are present Many species live here, and many more could if protected from competition. Species diversity here is high.
TROPHIC LEVELS
Trophic levels are basically feeding levels. Each ecosystem contains some members, autotrophs that bring energy into the system. Photosynthesis is by far the dominant method, accounting for virtually all energy input. Green vascular plants are most important, but algae and cyanobacteria carry on about one third of all photosynthesis worldwide, and lichens and nonvascular plants are important in cold, high latitudes and altitudes. Chemo- synthetic bacteria bring chemical energy into ecosystems, and although this is minor now, it was the only method before photosynthesis evolved (Fig. 8).
Autotrophs are known as primary producers for obvious reasons, and they are the first step of any food web. They are the energy and nutrient supply (food) for the herbivores, which constitute the primary consumers (sometimes called secondary producers). Herbivores are preyed on by carnivores, the secondary consumers. Omnivores exist at both trophic levels. Decomposers such as fungi and bacteria break down the remains of all types of organisms, even those of other decomposers (Fig. 9).

FIGURE 8: Recently, colonies of tube worms and clams have been discovered on the deep sea floor near hydrothermal vents, areas similar to Yellowstone's geysers, where hot, mineral-rich water erupts into the ocean. The water contains large amounts of hydrogen sulfide, which is oxidized by chemotrophic bacteria. However, the bacteria occur symbiotically within the tissues of the tube worms and clams, and they leak carbohydrates to their symbiotic hosts. Basically, the bacteria act like chemosynthetic "chloroplasts." A further exotic metabolism is that the blood of tube worms is able to carry hydrogen sulfide to the bacteria in their tissues, even though hydrogen sulfide is highly toxic to most animals. This is an unusual ecosystem that is not dependent on sunlight and in which plants are not involved at all. (]. Frederick Grassle/Woods Hole Oceanographic Institution)
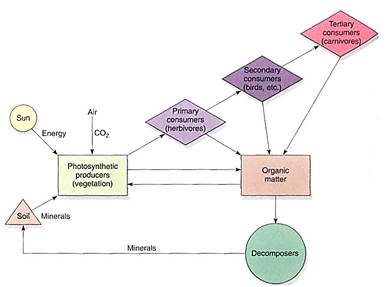
FIGURE 9: Energy and materials flow through the ecosystem as a network rather than in a straight line. Each level contributes to the decomposers (they even decompose other decomposers), and omnivores eat both plants and other animals. The primary producers never draw energy from the other stages, but many depend on animals and decomposers to return essential elements to the soil. At each step, respiration returns most of the carbon back to the air, and most of the energy is liberated as heat.
As plants photosynthesize, energy and carbon compounds enter the ecosystem. As the plants are eaten, the energy and the carbon compounds move to the herbivore trophic level, then to the carnivore trophic level, and finally on to the decomposers. This is referred to as the energy flow and the carbon flow of the ecosystem. At each step much of the food is used in respiration, resulting in the production of ATP, heat, carbon dioxide, and water. The carbon dioxide is released back to the atmosphere, where it can be used in photosynthesis again; the energy temporarily contributes to warming the planet, but it is ultimately radiated into space. The portion of the food that is not respired is available for growth and reproduction. As a very rough approximation, about 90% of an animal's food is respired and f 0% is retained as growth or gametes. Therefore, when herbivores eat 1000 kg of grass, 900 kg are respired and 100 kg become herbivore tissues. Similarly, a carnivore that eats the 100 kg of herbivore would only gain 10 kg, and after dying would support just 1 kg of decomposers.
Along with the flow of energy and carbon, minerals flow through the ecosystem. Plants absorb minerals from soil and incorporate them into their bodies as amino acids, ATP, coenzymes, and so on, and these are consumed by herbivores. Many of these compounds are broken down by animals, and the minerals are lost as waste. After organisms die, the minerals that remain in their bodies are released by decomposers and become part of the soil again. In addition, minerals can be completely lost from an ecosystem as they are carried away by rainwater and streams, especially if erosion is occurring or as cities dump mineral-rich human waste into rivers. Such minerals are carried to the oceans and become part of marine ecosystems. Many algae, protists, and marine animals have bodies with heavy, mineral-rich shells and bones; as the organisms die, the bodies sink to the depths of the ocean and are unavailable to living organisms of the land or ocean surface.
The movement of energy and biomass from one trophic level to the next is often represented as a pyramid of energy or a pyramid of biomass (Fig. 26.37). Such pyramids are useful for illustrating these principles in textbooks, but in reality it is extremely difficult to measure all the biomass of the primary producers of even a simple ecosystem. It is almost impossible to measure it accurately in a forest ecosystem, and it is impossible to measure the biomass or energy content of the fungi and bacteria that are acting as decomposers. Pyramids of numbers are more easily obtained; often there are fewer individuals in each higher trophic level, but of course millions of aphids can live on one tree.
Pyramids of biomass and numbers have become important as a means of illustrating the impact of the human introduction of herbicides, pesticides, and toxic wastes. Although some of these are biodegradable, many are not and they persist unchanged and toxic in the environment. Imagine a toxic herbicide applied at the low rate of one part per million—1 gm per 1000 kg of pasture grass. Herbivores will eat the 1000 kg of plants, and the pyramid of biomass shows that they will respire 900 kg away, but the 1 gm of herbicide is not oxidized or excreted. Instead it accumulates in the animals, now at a concentration of 1 gm per 100 kg. This is ten times stronger than the application rate. As carnivores eat the herbivores, the concentration rises to 1 gm per 10 kg, now 100 times stronger. Human metabolism is no different; these chemicals move through the food web to us as well. The toxic materials are not distributed uniformly within the bodies of animals and humans but become concentrated in our livers and kidneys as well as in fatty tissues such as our brain and spinal cord.
The role of decomposers in the ecosystem is vitally important; decomposers are so ubiquitous as well as microscopic that we take them for granted. If there were no decomposers in nature, plants could grow only as rapidly as weathering breaks down rock; as soon as minerals were released, they would be absorbed and locked into a plant and would then move through the food web. When the organisms died, the minerals would remain locked in the non-decomposing body. Sterilized (canned) food does not decay even though moist and kept at room temperature because no decomposers are present. Only by the physical and chemical weathering of rock and dead bodies could plant growth continue. But with decomposers present, minerals are recycled rapidly; decomposers release the minerals to the environment through their membranes, and their own bodies are so small and delicate that after death they are broken down immediately by other decomposers or by inorganic chemical weathering.
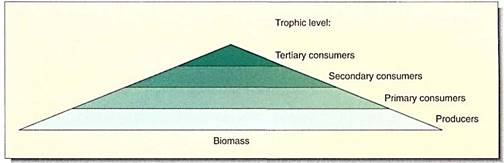
FIGURE 10: A pyramid such as this can represent the number of organisms at each level, the weight of all the individuals at each level, the respiration at each level, and so on. Such analysis can be difficult to do, but these relationships are extremely important. If a national park is established to preserve a species of wolf or mountain lion, it must be known how extensive its food web is and how large an area must be set aside to ensure an adequate number of primary consumers.
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















