























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Other Aspects of Inheritance
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 19-10-2016
19-10-2016
 2852
2852




+
-
20
Other Aspects of Inheritance
MATERNAL INHERITANCE
In the crosses described above, the alleles of both parents are transmitted equally to the progeny, a situation called biparental inheritance. All genes in the nucleus undergo biparental inheritance. An unexpected feature of both plants and animals is that during fertilization, the sperm cell loses most of its cytoplasm and only the sperm nucleus enters the egg. Consequently, the zygote obtains all its plastid and mitochondrion genomes from the maternal parent; this is uniparental inheritance, more specifically maternal inheritance.
Mitochondrion genetics are difficult to study for several reasons. First, each cell has many mitochondria, up to hundreds, and each mitochondrion has several circles of DNA. Each cell has multiple copies of each mitochondrion gene and may have received diverse types of alleles from its maternal parent. Simple mendelian ratios are not produced. Second, the presence of many of the mitochondrion DNA mutants does not result in easily recognizable phenotypes. The plants usually simply grow somewhat more slowly, or, if the mutation has a severe effect, they may die as embryos. Consequently, the presence of mitochondrion mutants may not be detectable by examination.
Plastid genetics are somewhat easier if alleles affecting chlorophyll synthesis are studied. Consider a plastid whose DNA has mutated so that the plastid can no longer produce any chlorophyll; it may be red or orange because the carotenoids are no longer masked by chlorophyll, or it may be white if no pigment at all is produced. If a zygote receives only plastids like these, the embryo and seedling will be red, orange, or white. Such seedlings are rare, but in large commercial nurseries that grow millions of seedlings, they appear from time to time. The plants can be kept alive artificially if they are grafted onto a normal plant that supplies them with carbohydrates (Fig. 1). Grafted plants can then be used for genetic experiments.

FIGURE 1:Plastid mutations in the cactus genus Gymnocalycium seem to be common; quite often totally red or orange seedlings appear in cactus nurseries. These lack chlorophyll but can be kept alive by grafting them onto a green cactus that supplies them with sugar. Many plants like Coleus and maples have bright red leaves; they do have chlorophyll, but it is masked by large amounts of other pigments.
When an achlorophyllous plant is crossed with a green, chlorophyll-bearing plant, the outcome depends on which plant is the pollen parent and which is the ovule parent. If the achlorophyllous parent provides the pollen, 100% of the zygotes will be green, because they receive all their plastids from the normal parent by way of the egg; all mutant plastids are destroyed during sperm differentiation and syngamy. But if the achlorophyllous plant is the ovule parent, all progeny also have mutant plastids and are achlorophyllous.
Plastid inheritance is also responsible for certain types of variegation in plants, the presence of spots or sectors that are white, red, or orange on a plant that is otherwise green (Fig. 2). Because plant cells have many plastids, only rarely do all the plastids of an egg have the same alleles unless the parent has only one type, as in the cactus of Figure 1622 More often, each cell has a mixture of plastid types. During cell division, some plastids move into each end of the dividing cell; usually random chance alone results in each daughter cell gelling some of both types of plastid (Fig. 16.23b). However, occasionally one daughter cell by chance receives only mutant plastids. It is white, red, or orange, and is able to survive because surrounding normal cells supply it with sugar. If this occurs ina leaf primordium or a young internode, the cell continues to grow and divide along with the surrounding cells, resulting in a patch of cells that lack chlorophyll (Fig. 16.23a). If it occurs in a very young leaf, the cells grow quite a lot and produce a large spot; if it happens in an older leaf that has reached almost full size, the patch is small.
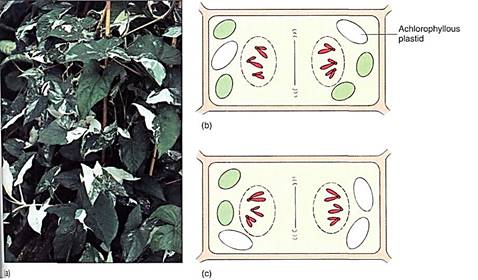
FIGURE 2: (a) Variegated plants of Syngonium podophyllum. Each green cell probably has a mixture of normal chloroplasts and plastids with a mutation that prevents chlorophyll synthesis. Cells in the white aea have only mutant plastids. (b and c) Cells usually have many plastids and mitochondria, so random distribution of the organelles is usually sufficient to ensure that both cells receive some Kb) If mutant plastids are present, then occasionally when cytokinesis occurs, one end of the cell may by chance have only mutant plastids (c). Most cells have only two sets of (nuclear) chromosomes, so all the elaborate mechanisms of mitosis are necessary if each daughter cell is to receive one complete set.
When variegation affects stems, the achlorophyllous patch may include an axillary bud. If so, the entire bud and the branch that grows from it are achlorophyllous. Any flowers it produces can be used for genetics experiments, providing either pollen or ovule& Experiments on both corn and morning glory have shown the achlorophyllous character to be maternally inherited; thus it is a feature of the plastid DNA as just described. Recombination DNA technology is now greatly accelerating our efforts to understand plastid and mitochondrion genetics. In both organelles the DNA is a circular double helix, one of its ends being attached to the other, and no histones are present. Plastids contain many circles, 200 in each spinach plastid and 1000 in each of those of wheat, so rather than being haploid or diploid, wheat plastids are 1000-ploid. Plastid DNA may constitute up to 21% of the total DNA content of a cell . In angiosperms, plastid DNA tends to be about 150 kbp long. In plants, mitochondrion DNA circles are very large, well over 200 kbp (up to 2400 kbp), more than ten times larger than mitochondrion DNA in animals.
Genomes of plastids contain genes for numerous enzymes as well as a complete set of plastid ribosomal RNAs and transfer RNAs. At present it is estimated that 80 to 100 enzymes and membrane proteins are coded by plastid DNA, and many of these are essential: the large subunit of RuBP carboxylase, components of the proton channel/ATP synthetase complex, cytochromes, and proteins that bind chlorophyll a (Fig. 3). Mitochondrion DNA also contains genes for rRNA and tRNA as well as those that code for proteins involved in ATP synthetase complexes, electron transport, and NADH dehydro-complex. In many cases, organelle genes must work in coordination with nuclear genes. The interaction of these three genomes within one cell is now an area of very active research.
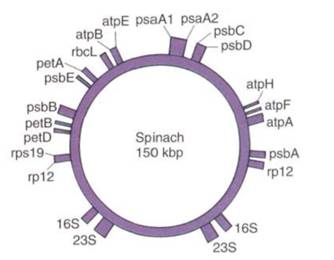
FIGURE 3:A map of the plastid DNA circle from spinach; the letters are the abbreviations for the names of the genes. For several species, the entire plastid circle has been sequenced so that we know the exact position of every nucleotide. (Based on data of J. D. Palmer)
LETHAL ALLELES
The phenotypic result of a mutation can vary in severity from almost undetectable to lethal, that is, its presence can kill the plant. Genetically inherited lethal mutations are most often recessive and are fatal only if present in the homozygous condition; a heterozygous plant has enough normal protein to survive. The reason for the recessive nature of m3 lethals is simple: A dominant lethal would kill the plant, even in the heterozygous condition, so it would rarely be passed on to any offspring.
The presence of a lethal allele can be difficult to detect if its effect occurs early. If it affects basic metabolic functions such as respiration, DNA replication, cellulose synthesis, or the structure of histones, plants that are homozygous for it probably die while still very young; in some instances the gametes die even before fertilization. The only seeds that complete development are heterozygotes or dominant homozygotes. This results in unusual genotype and phenotype ratios. A selling of a heterozygote for a lethal condition, Ll, produces zygotes in the typical 1 LL:2 Ll : 1 ll genotype ratio, but the ll zygotes die. The seeds and plants that live are 1 LL: 2 Ll genotypically; such a ratio should always lead one to suspect a recessive lethal allele.
Plants have an alternation of heteromorphic generations, the diploid sporophytes alternating with the haploid gametophytes. In gametophytes, each gene is present only as a single allele. No homologous chromosomes occur, and recessive and lethal alleles are not masked by the presence of a dominant homologous allele. In animals, this haploid phase is restricted to the sperm and egg, which are simple and have many inactive genes. As a result, numerous genes with deleterious mutations are not active during the brief haploid condition; they are turned on only during growth and differentiation from a zygote to an adult, when their effect is mitigated by their diploid condition. In plants, however, the haploid phase consists of at least a pollen grain with three cells and a megagametophyte with seven cells. This phase is still quite simple and most genes are inactive, but in both gametophytes many types of central metabolism are occurring, and any highly deleterious allele causes that gametophyte to develop poorly or not at all. If the gametophyte dies without producing a sperm or an egg, the allele is eliminated.
MULTIPLE SETS OF CHROMOSOMES
Whereas most animals have diploid body cells and haploid gametes, plants are much more diverse. Rarely, a spore is formed without undergoing meiosis, resulting in diploid game- tophytes and diploid gametes. If these fertilize a normal haploid gamete, a triploid zygotes results. Although this would be instantly lethal in almost all animals, plants tolerate it well and a triploid sporophyte develops from the zygote. However, triploids are sterile because they cannot undergo pairing of homologous chromosomes during prophase I; chromosomes come together in threes and the rest of meiosis is aberrant.
Occasionally, cells fail to undergo mitosis after S phase DNA replication, and the dl becomes tetraploid (or diploid if it happens in a gametophyte). Tetraploid cells are usually perfectly healthy; if they produce a part of the plant that initiates flowers, diploid pollen and eggs are produced, again raising the possibility of triploid zygotes if fertilized by haploid gametes. These processes can occur in virtually every conceivable combination, and plants that are 3n (three sets of chromosomes, triploid), 4n, 5n, 6n, ... up to several hundred n are common. All plants with more than two sets of chromosomes are polyploid. All plants with an even ploidy level can undergo meiosis and are fertile; all ad ploidy levels are sterile like triploids.
Change in chromosome number per nucleus in plants can also come about by non- disjunction: During the second division of meiosis, the two chromatids may remain together (fail to disjoin), so one daughter cell receives both copies of the chromosome (the two chromatids) whereas the other receives none. The cell with the extra chromosome probably survives quite well and forms a functional sperm or egg cell. If this gamete, which is diploid for one chromosome, is involved in fertilization, the new zygote will be triploid for that chromosome. It received a normal, haploid chromosome set from one parent and a haploid set plus the extra chromosome from the parent in which nondisjunction occurred. In plants, this condition is tolerated often, and only by studying the karyotype can it be discovered. In animals, this is almost invariably fatal. The human congenital disease Down syndrome is caused by nondisjunction of the very small chromosome 21. Down syndrome individuals are triploid for just a few genes yet have severely disrupted metabolism. Fetuses triploid for longer chromosomes abort spontaneously.
Plants with even ploidy levels grow and reproduce successfully, often more vigorously than diploid individuals of their species. Initially, these have complex genetics. For example, a tetraploid can have any of the following genotypes lor red/white flower color: RRRR, RRRr, RRrr, Rrrr, or rrrr. Its gametes can be RR, Rr, or rr. Notice that polyploids have more copies than needed for every gene. In all diploid plants, even heterozygotes with only one functional allele are usually healthy. For nonessential genes, a complete absence of functional alleles may not be harmful, such as when rr produces white flower color. conse- quently, the two or three extra copies of each gene in a tetraploid plant are surplus DNA. Mutations that completely incapacitate one of the four alleles of a tetraploid usually are not deleterious, as one, two, or three functional alleles remain in every nucleus. For polyploid plants, deleterious mutations tend to have little effect on phenotype, so they do not affect survival as much as they would in a haploid cell or a diploid plant. As a result, deleterious mutations are not eliminated quickly by natural selection, and the extra copies of a gene may rapidly become nonfunctional; before many generations have passed, polyploids act like diploids, their "extra" copies having mutated. Almost half of all flowering plant species are actually polyploids; those that now appear to be perfectly normal diploids probably underwent conversion to the polyploid condition so long ago that all their extra alleles have mutated extensively and can no longer be considered alleles of those that are still functional. Nondisjunction, insertions, and deletions may have changed the numbers and sizes of the chromosomes so much that the four originally homologous chromosomes now appear as just two different pairs. Conversion back to the diploid state is an uneven process, and some species act like diploids for certain genes but tetraploids for others.
What do the extra alleles mutate into? At first they may simply be alleles that produce defective proteins. Mutations in the promoter regions may prevent them from ever being activated, so they turn into "junk," acting as nothing more than spacer DNA. Deletions, inversions, and movements by transposons may break them up. DNA sequencing has now revealed that there are large families of genes in which one gene codes for a useful protein while the other genes have nucleotide sequences that are obviously related to the useful gene but are inactive and apparently code for nothing.
This extra DNA is actually extremely valuable—it is the raw material for the evolution of new genes. Some may mutate into genes that produce enzymes almost identical to those being coded by the original form of the allele. The original enzyme may work best at low temperature, whereas the new form may have a higher optimum temperature. The plant can now produce two types of enzymes and function well in both warm and cool days, or the species may be able to extend its range, the new enzyme allowing it to survive in hot deserts and the original enzyme allowing it to live in its original habitat (Fig.4).

FIGURE 4:Once a plant becomes tetraploid, mutations in the extra copies of a gene are tolerable as long as (1) at least one allele is left functional, and (2) the extra copies do not mutate into a lethal allele. In this example, two chromosomes have changed morphology, becoming shorter; they could have become longer by picking up pieces of DNA by unequal crossing-over. Several copies have mutated into spacers and some into junk that still has much sequence homology with functional alleles. Others have evolved into new alleles of the original genes (d3 to d5; a2 to a3; ]3 to 4), and some have evolved into entirely new genes (a1 to g1; d4 to h1).
Mutations in the promoter region instead may allow the structural portion to produce the same protein as the original gene but at a different time or place in response to a different chemical messenger.
In other cases, mutations in the extra DNA may result in totally new genes that produce proteins whose function is not at all related to that of the original gene. Mosses, ferns, conifers, and flowering plants have evolved from green algae. Because algae have no genes for flower color, lignin synthesis, and so on, the evolution of these species has involved the evolution of whole new metabolic pathways. All of these genes had to arise by the gradual mutation of surplus alleles into nucleotide sequences that code for useful proteins.
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















