























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Types of Evidence Used For Taxonomic Analysis
 المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
المؤلف:
AN INTRODUCTION TO PLANT BIOLOGY-1998
 المصدر:
JAMES D. MAUSETH
المصدر:
JAMES D. MAUSETH
 الجزء والصفحة:
الجزء والصفحة:
 17-10-2016
17-10-2016
 3968
3968




+
-
20
Types of Evidence Used For Taxonomic Analysis
HOMOLOGY AND ANALOGY
The study of plant phylogeny centers on examining the similarity of one species to others. A species evolves into two species as different populations accumulate distinct alleles; even when enough divergence has occurred to create separate species, the two still resemble each other strongly. As they continue to evolve each acquires its own mutations, and because they cannot interbreed they cannot share the new alleles (Fig. 1). Thus, fay differ from each other more as time passes. Distantly related plants have been on separate lines of evolution for millions of years, time enough for so many mutations to accumulate that they resemble each other only slightly.
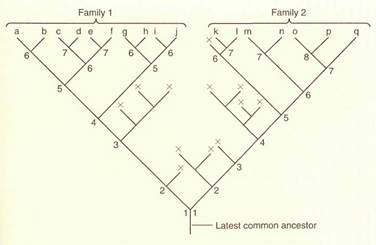
FIGURE 18.6: In this hypolhetical evolutionary history, two lines diverged long ago, then many of the early species became extinct (lines marked with x), and only some continued on, speciating into two separate families. In family 1 all extant species have at least six speciation events in their history since the latest common ancestor shared with family 2. Family 2 has undergone slightly more rapid evolution: The minimum number of branch points is seven, and two species have eight. Thus, since these two lines of evolution diverged, many changes have occurred in each, so they are quite distinct. In reality, there are about 400,000 species of vascular plants with unknown hundreds of thousands or millions of extinct species, so the true phylogeny is gigantic. The latest common ancestor for vascular plants probably occurred about 420 million years ago.
Taxonomy and systematics consist of studying various species and examining their degrees of similarity, but this study is complicated by the fact that plants can resemble each other for two distinct reasons: (i) they have descended from a common ancestor, or (2) they have undergone convergent evolution. Features similar to each other because they have descended from a common ancestral feature are homologous features. For instance, almost all members of the anthurium family are easily recognizable because of their spathe and spadix inflorescence (Fig. 2). All members have these structures because all have inherited their inflorescence genes from a common ancestor. The spathe and spadix are homologous in all Anthurium species. Homologous features are the ones critically important for making taxonomic comparisons and the only ones that can be used to conclude that species are related.

FIGURE 2 :Members of the family Araceae (the aroids, Jack- in-the-pulpit) all have a spathe (the collar-like sheath) and spadix (the central column), as in this Amorphophallus (a) and Anthurium (b). In all extant species, the genes that control the formation of these structures are descendents of the genes of the original, ancestral species. The genes that exist today have resulted from thousands of rounds of DNA replication during plant growth, crossing-over, syngamy, and then more rounds of DNA replication as the zygote grows. This has been repeated millions of times since spathes and spadices first evolved millions of years ago. During all this, mutations and natural selection have led to obvious varieties of spathes and spadices, but the basic relatedness, the homology, is still obvious.
The second cause of resemblance, convergent evolution, results when two distinct evolutionary lines of plants respond to similar environments and selection pressures. Under these conditions, natural selection may favor mutations in each line that result in similar phenotypes. Features like this are analogous features and should never be used to conclude that plants are closely related. . Both occur in deserts where a succulent water-storing body is advantageous and spines are selectively advantageous by deterring animals from eating the plants to get the water. The two families are not considered closely related on the basis of spines and succulence, however, because these features do not share a common ancestry; they converged evolutionarily because of similar selection pressure.
Determining whether a similarity is due to homology (common ancestry) or analogy (convergent evolution) can be extremely difficult. In the case of spines, it is easy: Cactus spines are modified leaves that are always smooth and never branched, occurring in clusters on an extremely short shoot. Euphorbia spines are modified shoots that branch and have small, scalelike leaves. Being modified shoots, they occur singly in the axil of a leaf, never in clusters. If we look beyond the analogous similarities, the dissimilar features are dramatically different; flowers of cacti are large and have many parts, whereas those of euphorbias are small and unisexual, either staminate or carpellate. The wood of cacti is parenchymatous and has few fibers whereas that of euphorbias is very fibrous and hard. MS important, the plants that most closely resemble the succulent euphorbias in important homologous features are the nonsucculent euphorbias of Africa. Succulent cacti resemble a completely different set of plants, nonsucculent cacti of the tropical regions of the Americas. Thus, the evolution of spines and succulence occurred independently in each group.
A corollary to the assumption that similar plants are closely related is the assumption that dissimilar plants must not be closely related. Studying lack of similarity can also be difficult because in some cases, a small genetic change results in dramatic phenotypic changes (Fig. 3). Mutations that affect production, distribution, and sensitivity to hormones result in large changes of the phenotype between two closely related species. Also, mutations that affect early stages of development such as the embryo or bud meristems can cause closely related species to look deceptively dissimilar. In many evolutionary lines, species of large woody trees have evolved into herbs; the main transition involved is the ability to flower early and then not survive a harsh winter. It must be a simple evolutionary process involving few genes because it has occurred many times. For example, nearly half the species of the Violaceae are herbs in the genus Viola—violets and pansies—but the early members of the family were woody trees, as are the members of the relictual, tropical genus Rinorea. Artificial selection by humans has resulted in one species that has dramatically different forms: Broccoli, brussels sprouts, cabbage, and cauliflower are all subspecies of a single species, Brassica oleracea.

FIGURE 3 :This small, leafless shrub (a, Acacia aphylla) may not appear at first glance to have much in common with this larger, leafy tree (b, Acacia drepanolobium), but careful examination reveals homologous similarities in many critical features of the flowers and stems which allow assignment to the same genus, Acacia. In this case, genes controlling flower and stem morphogenesis have changed little during evolution, whereas genes controlling elongation growth of stems, formation of leaf primordia, and formation of bark have all changed dramatically.
For taxonomic studies, a mosaic of evidence is available—some valid and useful, some misleading. Taxonomists attempt to look at as many features as possible on the assumption that misleading evidence will be outweighed by valid characters. Also, certain features are considered more significant; those that result from numerous metabolic interactions and the influence of many genes tend to change more slowly than those with a simple metabolism controlled by just one or two genes. Loss of features, such as sepals and petals in wind-pollinated flowers, is often rapid because deleterious mutations occur commonly; without the natural selection of insect pollination to eliminate these mutations, they are retained and cause a loss of perianth. Once those genes have become so mutated that a flower lacks sepals and petals, the evolution of new ones would be slow and difficult.
At present, taxonomists study virtually every aspect of plants using a wide variety of tools. Simple observation of major parts is still important, but scanning electron microscopy to study hairs, stomata, cuticle, and waxes is also used. Internal structure is studied with both light and transmission electron microscopy; the nature of plastids in the phloem has been found to be an extremely valuable character . Various aspects of metabolism are important, ranging from the types of pigments in flowers to the presence of CAM and C4 metabolism and specialized defensive, antipredator compounds.
DNA sequencing is a new tool for analyzing evolutionary relationships (Fig. 4). Two plants are considered separate species only if they differ in significant, heritable ways; therefore, the sequence of nucleotides in the DNA of each species must differ from that of all other species. When plant phenotypes arc studied, the actual objective is to determine differences in genotype. In the past, this could be done only by examining phenotypic features and inferring genotype differences if they could not be related to direct environmental effects. Now DNA can be examined directly and mutations can be identified, even if they do not cause any detectable change in phenotype.

FIGURE 4 : Chloroplast DNA was extracted from plants of three species of Machaeranthera (in the sunflower family), then cut into fragments with the restriction endonuclease ClaI. The restriction map of M. canescens plastid DNA is almost identical to that of M. bigelovii, which has every one of the 56 restriction sites present in M. canescens plus one more (blue to left of middle). M. riparia is rather distinct: It lacks only one of the sites common to the other two species (near its right end"1, but it has five that do not occur in the other species (red). Black—present in all three species; blue- present in two species; red—present in only one species.
Like traditional taxonomic data, DNA sequencing data have their own ambiguities and limitations. Any two species, even those closely related, must differ at many parts (nucleotide pairs) of their DNA. Simply counting the number of sequence differences is not enough, because even individuals of a very uniform, homogeneous species may have hidden mutations that do not affect the phenotype. Furthermore, any two species in the same family probably share the great majority of their DNA sequences in identical form, with only a few genes mutated. How many differences are needed to conclude that two genomes represent two distinct species? There is no simple answer. If it were necessary to examine the entire genome to find the few points of difference, the problem would be insurmountable. A quick glance instantly reveals that two anthuriums are more closely related to each other than to a geranium, cactus, or euphorbia, but demonstrating the same thing using DNA techniques would take many weeks and a great deal of money.
 الاكثر قراءة في مواضيع عامة في علم النبات
الاكثر قراءة في مواضيع عامة في علم النبات
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















