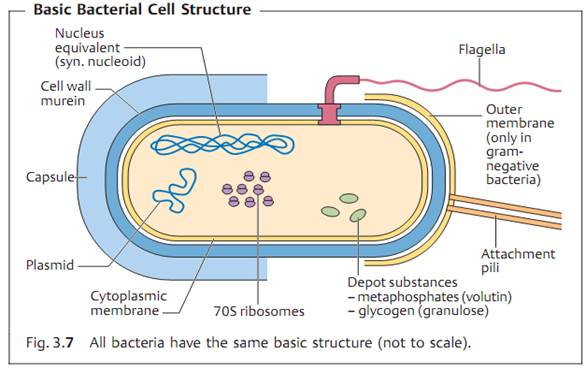
Fine Structures of Bacteria
Nucleoid (Nucleus Equivalent) and Plasmids
The “cellular nucleus” in prokaryotes consists of a tangle of double-stranded DNA, not surrounded by a membrane and localized in the cytoplasm (Fig. 3.5). In E. coli (and probably in all bacteria), it takes the form of a single circular molecule of DNA. The genome of E. coli comprises 4.63 x 106 base pairs (bp) that code for 4288 different proteins. The genomic sequence of many bacteria is known.
The plasmids are nonessential genetic structures. These circular, twisted DNA molecules are 100-1000 x smaller than the nucleoid genome structure and reproduce autonomously (Fig. 3.6). The plasmids of human pathogen bacteria often bear important genes determining the phenotype of their cells (resistance genes, virulence genes).


Cytoplasm
The cytoplasm contains a large number of solute low- and high-molecular- weight substances, RNA and approximately 20 000 ribosomes per cell. Bacteria have 70S ribosomes comprising 30S and 50S subunits. Bacterial ribosomes function as the organelles for protein synthesis. The cytoplasm is also frequently used to store reserve substances (glycogen depots, polymerized metaphosphates, lipids).
The Cytoplasmic Membrane
This elementary membrane, also known as the plasma membrane, is typical of living cells. It is basically a double layer of phospholipids with numerous proteins integrated into its structure. The most important of these membrane proteins are permeases, enzymes for the biosynthesis of the cell wall, transfer proteins for secretion of extracellular proteins, sensor or signal proteins, and respiratory chain enzymes.
In electron microscopic images of Gram-positive bacteria, the mesosomes appear as structures bound to the membrane. How they function and what role they play remain to be clarified. They may be no more than artifacts.
Cell Wall
The tasks of the complex bacterial cell wall are to protect the protoplasts from external noxae, to withstand and maintain the osmotic pressure gradient between the cell interior and the extracellular environment (with internal pressures as high as 500-2000 kPa), to give the cell its outer form and to facilitate communication with its surroundings.
Murein (syn. peptidoglycan). The most important structural element of the wall is murein, a netlike polymer material surrounding the entire cell (sacculus). It is made up of polysaccharide chains crosslinked by peptides (Figs. 3.8 and 3.9).
The cell wall of Gram-positive bacteria (Fig. 3.10). The murein sacculus may consist of as many as 40 layers (15-80 nm thick) and account for as much as 30% of the dry mass of the cell wall. The membrane lipoteichoic acids are anchored in the cytoplasmic membrane, whereas the cell wall teichoic acids are covalently coupled to the murein. The physiological role of the teichoic acids is not known in detail; possibly they regulate the activity of the auto- lysins that steer growth and transverse fission processes in the cell.



Within the macroorganism, teichoic acids can activate the alternative complement pathway and stimulate macrophages to secrete cytokines. Examples of cell wall-associated proteins are protein A, the clumping factor, and the fibronec- tin-binding protein of Staphylococcus aureus or the M protein of Streptococcus pyogenes. Cell wall anchor regions in these proteins extending far beyond the murein are bound covalently to its peptide components. Cell wall-associated proteins frequently function as pathogenicity determinants (specific adherence; phagocyte protection).
The cell wall of Gram-negative bacteria. Here, the murein is only about 2 nm thick and contributes up to 10% of the dry cell wall mass (Fig. 3.11). The outer membrane is the salient structural element. It contains numerous proteins (50% by mass) as well as the medically critical lipopolysaccharide.
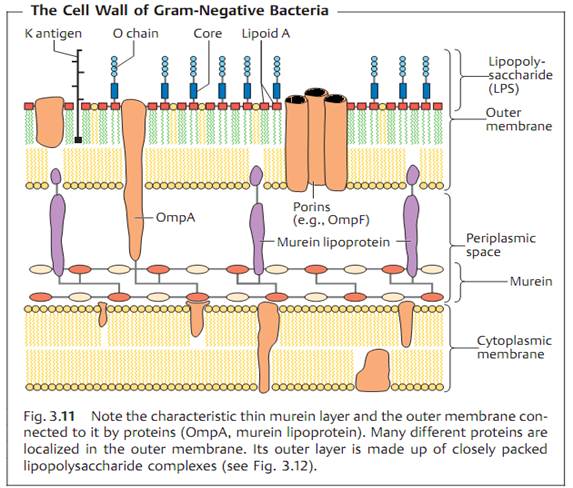
Outer membrane proteins
- OmpA (outer membrane protein A) and the murein lipoprotein form a bond between outer membrane and murein.
- Porins, proteins that form pores in the outer membrane, allow passage of hydrophilic, low-molecular-weight substances into the periplasmic space.
- Outer membrane-associated proteins constitute specific structures that enable bacteria to attach to host cell receptors.
- A number of Omps are transport proteins. Examples include the LamB proteins for maltose transport and FepA for transport of the siderophore ferric (Fe3+) enterochelin in E. coli .
Lipopolysaccharide (LPS). This molecular complex, also known as endotoxin, is comprised of the lipoid A, the core polysaccharide, and the O-specific polysaccharide chain (Fig. 3.12).

Lipoid A is responsible for the toxic effect. As a free substance, or bound up in the LPS complex, it stimulates—by binding together with the LPS binding protein (LBP) to the CD14 receptor of macrophages—the formation and secretion of cytokines that determine clinical endotoxin symptomatology. Interleukin 1 (IL-1) and tumor necrosis factor (TNF) induce an increased synthesis of prostaglandin E2 in the hypothalamus, thus setting the “ther-mostat” in the temperature control center higher, resulting in fever. Other direct and indirect endotoxin effects include granulopoiesis stimulation, aggregation and degeneration of thrombocytes, intravasal coagulation due to factor VII activation, a drop in blood pressure, and cachexia. LPS can also activate the alternative complement pathway. Release of large amounts of endotoxin can lead to septic (endotoxin) shock. Endotoxin is not inactivated by vapor sterilization. Therefore, the parent materials used in production of parenteral pharmaceuticals must be free of endotoxins (pyrogens).
The O-specific polysaccharide chain is the so-called O antigen, the fine chemical structure of which results in a large number of antigenic variants useful in bacterial typing (e.g., detailed differentiation of salmonella types) .
L-forms (L = Lister Institute). The L-forms are bacteria with murein defects, e.g., resulting from the effects of betalactam antibiotics. L-forms are highly unstable when subjected to osmotic influences. They are totally resistant to betalactams, which block the biosynthesis of murein. The clinical significance of the L-forms is not clear. They may revert to the normal bacterial form when betalactam therapy is discontinued, resulting in a relapse.
Capsule
Many pathogenic bacteria make use of extracellular enzymes to synthesize a polymer that forms a layer around the cell: the capsule. The capsule protects bacterial cells from phagocytosis. The capsule of most bacteria consists of a polysaccharide. The bacteria of a single species can be classified in different capsular serovars (or serotypes) based on the fine chemical structure of this polysaccharide.
Flagella
Flagella give bacteria the ability to move about actively. The flagella (singular flagellum) are made up of a class of linear proteins called flagellins. Flagel-

lated bacteria are described as monotrichous, lophotrichous, or peritrichous, depending on how the flagella are arranged . The basal body traverses the cell wall and cytoplasmic membrane to anchor the flagellum (3.13) and enables it to whirl about its axis like a propeller. In Enterobacteriaceae, the flagellar antigens are called H antigens. Together with the O antigens, they are used to classify bacteria in serovars.
Attachment Pili (Fimbriae), Conjugation Pili
Many Gram-negative bacteria possess thin microfibrils made of proteins (0.1¬1.5 nm thick, 4-8 nm long), the attachment pili. They are anchored in the outer membrane of the cell wall and extend radially from the surface. Using these structures, bacteria are capable of specific attachment to host cell re¬ceptors (ligand—receptor, key—keyhole).
The conjugation pili (syn. sex pili) in Gram-negative bacteria are required for the process of conjugation and thus for transfer of conjugative plasmids .

Biofilm
A bacterial biofilm is a structured community of bacterial cells embedded in a self-produced polymer matrix and attached to either an inert surface or living tissue. Such films can develop considerable thickness (mm). The bacteria located deep within such a biofilm structure are effectively isolated from immune system cells, antibodies, and antibiotics. The polymers they secrete are frequently glycosides, from which the term glycocalyx (glycoside cup) for the matrix is derived.
Bacterial Spores
Bacterial spores (endospores) are purely dormant life forms. Their development from bacterial cells in a “vegetative” state does not involve assimilation of additional external nutrients. They are spherical to oval in shape and are characterized by a thick spore wall and a high level of resistance to chemical and physical noxae. Among human pathogen bacteria, only the genera Clostridium and Bacillus produce spores. The heat resistance of these spores is their most important quality from a medical point of view, since heat sterilization procedures require very high temperatures to kill them effectively. Potential contributing factors to spore heat resistance include their thick wall structures, the dehydration of the spore, and crosslinking of the proteins by the calcium salt of pyridine-2,6-dicarboxylic acid, both of which render protein denaturing difficult. When a spore's milieu once again provides favorable conditions (nutrient medium, temperature, osmotic pressure, etc.) it returns to the vegetative state in which spore-forming bacteria can reproduce.





















