























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

The Innate Response Utilizes Conserved Recognition Molecules and Signaling Pathways
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 28-4-2021
28-4-2021
 2565
2565




+
-
20
The Innate Response Utilizes Conserved Recognition Molecules and Signaling Pathways
KEY CONCEPTS
- Innate immunity is triggered by pattern recognition receptors (PRRs), which recognize highly conserved microbe-associated molecular patterns (MAMPs) found in bacteria, viruses, and other infectious agents.
- Toll-like receptors (TLRs) are evolutionarily conserved and can direct both innate and adaptive immune responses.
- Natural antibodies are produced by adaptive immune cells (B lymphocytes) but mediate innate immunity.
As the first line of defense against microbial pathogens, innate immunity is activated upon recognition of certain predefined patterns in microorganisms by immune cell–associated pattern recognition receptors (PRRs). Most PRR ligands are conserved among microorganisms and are not found in higher eukaryotes, thereby allowing the immune system to quickly distinguish dangerous nonself from self. These microbe-associated molecular patterns (MAMPs) are synthesized by several sequential microbial enzyme reactions and, therefore, mutate more slowly than protein antigens (TABLE .1). Notably, nonpathogenic bacteria, such as commensal bacteria residing in the gut, also display conserved MAMPs.
TABLE .1 Innate immunity: A summary of MAMPs and PRRs.The Innate Response Utilizes
Conserved Recognition Molecules and Signaling Pathways
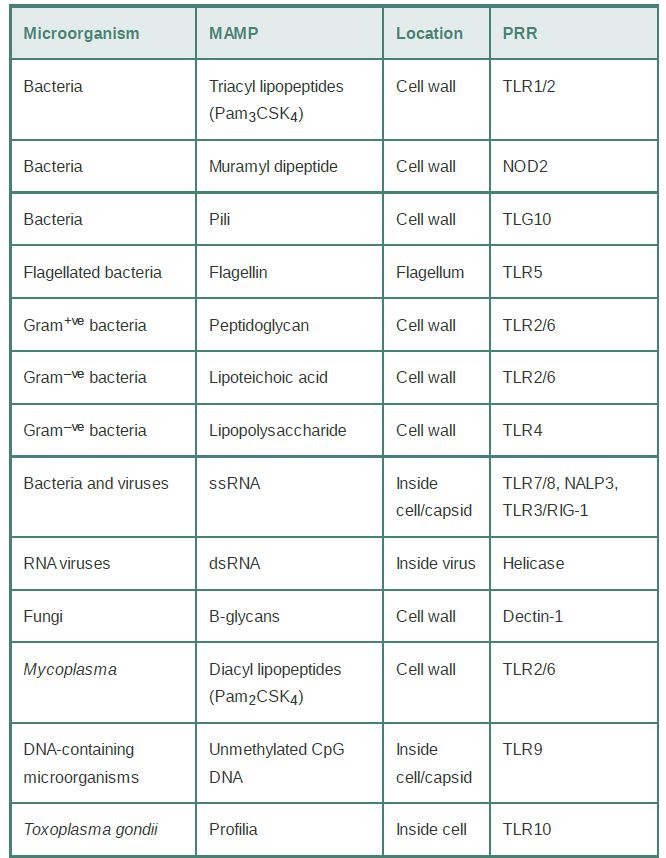
An important type of PRR is the Toll-like receptors (TLRs). TLR4 recognizes Gram-negative bacterial lipopolysaccharide (LPS), a well-known MAMP; TLR1 and TLR2 recognize lipoteichoic acid from Gram-positive bacteria and peptidoglycans; and TLR5 recognizes bacterial flagellin. These TLRs are expressed on the surface of immune cells. TLRs that recognize nucleic acid variants are normally associated with viruses, such as single-stranded RNA (TLR3), double-stranded RNA (TLR7 and TLR8), or certain unmethylated CpG DNA. TLR9 is localized in the cytoplasm. Upon sensing their ligands, TLRs rapidly activate innate immune responses by triggering activation of transcription factors for inflammatory gene expression. Notably, some TLRs also serve as sensors for selective environmental cues. For example, TLR4 recognizes nickel and mediates allergy to this metal.
Retinoic acid–inducible gene 1 (RIG-I) and RIG-I-like receptors (RLRs) are RNA sensors. RIG-I is activated by the 5′-triphosphate (5′-PPP) moiety of uncapped double-stranded RNA (dsRNA) or single-stranded RNA (ssRNA) of relatively short lengths, as typically found in replication intermediates of RNA viruses. This distinguishes viral RNA from usually capped eukaryotic mRNA. The RNA binding is mediated by the central RNA helicase DEAD box motifs and the C-terminal domain of RIG-I. The N-terminal caspase activation and recruitment domain (CARD) mediates the activation of downstream pathways to induce type I interferons for antiviral responses. Among other known members of the RLR family, MDA5 binds to 5′-PPP and triggers antiviral immunity, and LGP2 can only bind RNA but does not activate downstream pathways due to the lack of a CARD domain, thereby playing mainly regulatory roles.
Cyclic GMP-AMP (cGAMP) synthase (cGAS) is a recently identified sensor for cytosolic DNA, as associated with DNA virus and retrovirus replication. Upon activation by DNA, cGAS mediates the synthesis of cGAMP, a second messenger signaling molecule that, through its 2′–5′ phosphodiester linkage, activates pathways for the induction of antiviral type I interferon responses. Intercellular transmission of cGAMP, through tight junctions or by virus particles that package cGAMP, also allows the spread of the response to bystander immune cells. A homolog of cGAS is the oligoadenylate synthase (OAS) family of proteins, which can sense dsRNA and mediate the synthesis of 2′,5′–linked oligonucleotides to trigger immunity.
Innate response pathways are widely conserved and are found in organisms ranging from flies to humans. As the first identified and most studied PRRs, TLRs are orthologs of the Drosophila protein Toll. Toll, in addition to orchestrating dorsal–ventral organization during development, mediates innate antimicrobial activities. It is triggered by Spatzle, an insect cytokine produced by a proteolytic cascade upon infection by fungi or Gram-positive bacteria to activate Dorsal-related immunity factor (DIF), which is related to the mammalian transcription factor NF-κB. DIF, in turn, promotes expression of genes encoding antifungal peptides, such as drosomycin, which kill their respective target organisms through membrane permeabilization (FIGURE .1). The antibacterial response in flies also relies on peptidoglycan recognition proteins (PGRPs), which have high affinities for bacterial peptidoglycans.
Such responses lead to production of bactericidal peptides in a manner dependent on DIF or Relish, another NF-κB–related transcription factor, in response to Gram-positive and Gramnegative bacteria, respectively.
The TLR pathway in vertebrates is parallel to the Toll pathway with several equivalent components. About 10 human homologs of the TLRs can activate several immune response genes. Once a TLR is activated by an MAMP (as contrasted to the cytokine Spätzle in insects) it undergoes conformational changes and interacts, through homo- and heterodimerization, with one or more of five known Toll/interleukin 1/resistance (TIR) domain–containing adapters. These include myeloid differentiation primary response gene 88 (MyD88) and TIR domain–containing adapter-inducing interferon-β (TRIF), which, in turn, relay the signal, eventually leading to the induction of transcription factors such as NF-κB, AP-1, and IRFs for specific gene expression (FIGURE 2). The downstream pathways of TLRs are more expanded and versatile in mammals, as compared to those in insects. Notably, plants also use proteins with a leucine-rich region (LRR), which is the MAMPbinding site in TLRs, to detect pathogens and activate a mitogenactivated protein kinase (MAPK) cascade for induction of diseaseresistance genes.
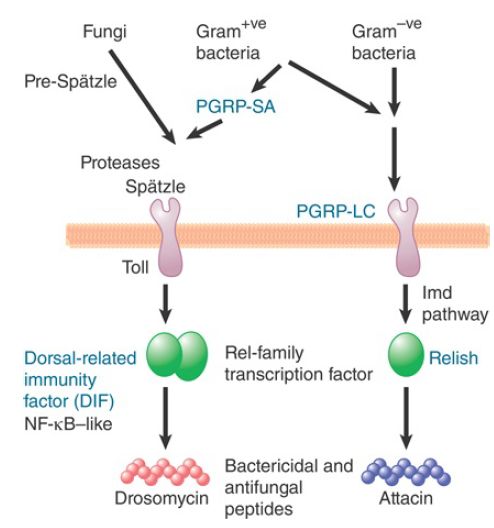
FIGURE .1 One of Drosophila’s innate immunity pathways is closely related to the mammalian pathway for activating NF-κB; the other has components related to those of apoptosis pathways.
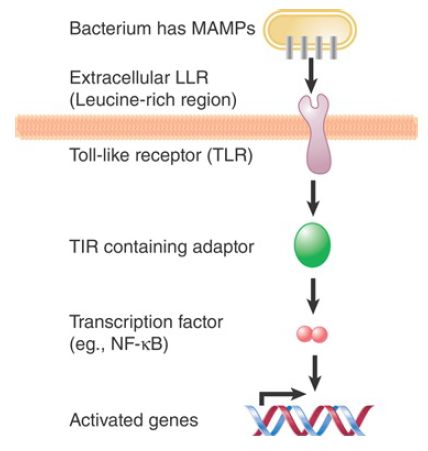
FIGURE .2 Innate immunity is triggered by MAMPs. In mammals, MAMPs cause the production of peptides that activate Toll-like receptors. The receptors lead to a pathway that activates a transcription factor for the Rel family. Target genes for this factor include bactericidal and antifungal peptides. The peptides act by permeabilizing the membrane of the pathogenic organism.
PRRs, particularly TLRs, are highly expressed in immune cells of the myeloid origin, such as neutrophils, macrophages, and DCs, which are capable of phagocytosing or killing pathogens directly, consistent with their innate immune functions. Several TLRs are also highly expressed in lymphocytes (i.e., B cells and selected T cell subsets).
In general, the innate response contains the first wave of invasion by pathogens, but cannot deal effectively with the later stages of virulent infections, which require the specificity and potency of the adaptive response. Innate and adaptive responses overlap and crosstalk, in that cells activated by the innate response subsequently participate in the adaptive response. This is exemplified by the B cell–intrinsic function of TLR signaling in adaptive immunity and the “innate” function of natural antibodies.
Natural antibodies are produced by B lymphocytes through the same DNA recombination process that generates BCRs and antibodies, in contrast to the aforementioned PRRs, which are
encoded by the germline. They are mainly IgM and are polyreactive (i.e., capable of binding multiple antigens). These antigens are often different in nature, such as phospholipids, polysaccharides, proteins, and nucleic acids, and are unlikely to share an identical epitope (which is the binding motif of an antibody). Rather, natural antibodies recognize foreign antigens possessing molecular structures that are different but that can equally fit the same natural antibody binding site—in this sense, natural polyreactive antibodies are also PRRs. This is exemplified by the ability of natural antibodies to bind appropriately spaced phosphate residues in the context of a variety of polynucleotides and phospholipids. Finally, many natural antibodies are “natural autoantibodies,” because they are produced in healthy individuals by B lymphocytes that show a moderate reactivity to a self-antigen and evade clonal deletion.
Natural polyreactive antibodies play an important role in early stages of infection, prior to the emergence of class-switched highly antigen–specific antibodies. They can also function as templates for the generation of high-affinity autoantibodies through somatic hypermutation.
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















