
Retroviruses May Transduce Cellular Sequences
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 24-4-2021
24-4-2021
 2599
2599
Retroviruses May Transduce Cellular Sequences
KEY CONCEPT
- Transforming retroviruses are generated by a recombination event in which a cellular RNA sequence replaces part of the retroviral RNA.
An interesting light on the viral life cycle is cast by the occurrence of transducing viruses, which are variants that have acquired cellular sequences in the form illustrated in FIGURE 1. Part of the viral sequence has been replaced by the v-onc gene. Protein synthesis generates a Gag-v-Onc protein instead of the usual Gag, Pol, and Env proteins. The resulting virus is replication defective; it cannot sustain an infective cycle by itself. It can, however, be perpetuated in the company of a helper virus that provides the missing viral functions.
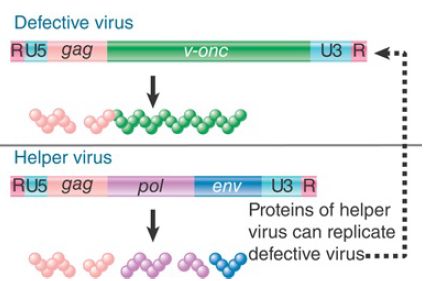
FIGURE 1.Replication-defective transforming viruses have a cellular sequence substituted for part of the viral sequence. The defective virus may replicate with the assistance of a helper virus that carries the wild-type functions.
Onc is an abbreviation for oncogenesis, the ability to transform cultured cells so that the usual regulation of growth is released to allow unrestricted division. Both viral and cellular onc genes may be responsible for creating tumorigenic cells.
A v-onc gene confers upon a virus the ability to transform a certain type of host cell. Loci with homologous sequences found in the host genome are called c-onc genes. How are the onc genes acquired by the retroviruses? A revealing feature is the discrepancy in the structures of c-onc and v-onc genes. The c-onc genes usually are interrupted by introns, whereas the v-onc genes are uninterrupted.
This suggests that the v-onc genes originate from spliced RNA copies of the c-onc genes. A model for the formation of transforming viruses is illustrated in FIGURE 2. A retrovirus has integrated near a c-onc gene. A deletion occurs to fuse the provirus to the c-onc gene; transcription then generates a joint RNA, which contains viral sequences at one end and cellular onc sequences at the other end. Splicing removes the introns in the cellular parts of the RNA. The RNA has the appropriate signals for packaging into the virion, which will be present if the cell also contains another intact copy of the provirus. At this point, some of the diploid virus particles may contain one fused RNA and one viral RNA.
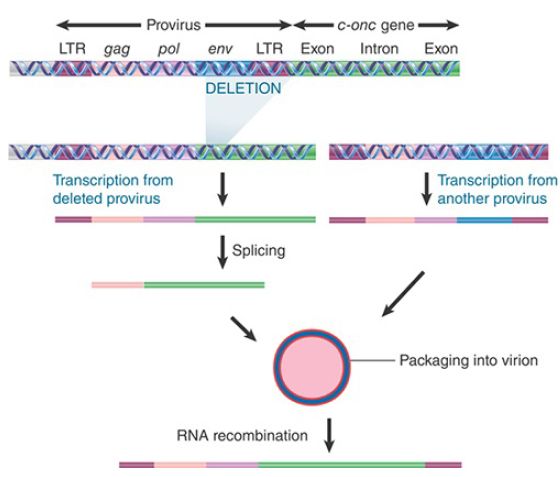
FIGURE 2. Replication-defective viruses may be generated through integration and deletion of a viral genome to generate a fused viral–cellular transcript that is packaged with a normal RNA genome. Nonhomologous recombination is necessary to generate the replication-defective transforming genome.
A recombination between these sequences could generate the transforming genome, in which the viral repeats are present at both ends. Recombination occurs by various means at a high frequency during the retroviral infective cycle. We do not know anything about its demands for homology in the substrates, but we assume that the nonhomologous reaction between a viral genome and the cellular part of the fused RNA proceeds by the same mechanisms responsible for viral recombination.
The common features of the entire retroviral class suggest that it may be derived from a single ancestor. This is supported by phylogenetic analysis of reverse transcriptases from a wide variety of retroelements, including both retrotransposons and retroviruses.
The fact that this class of elements has features common to both DNA-type transposons (integrase/transposase) and non-LTR retroposons (reverse transcriptase) has led to the suggestion that LTR retrotransposons arose as a consequence of a fusion between these two, more ancient element classes. Other functions, such as Env proteins and transforming genes, would have been incorporated later. (There is no reason to suppose that the mechanism is involved in acquisition of env and onc genes; viruses carrying these genes may have a selective advantage, though.)
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة





















