























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

How Did Interrupted Genes Evolve
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 16-3-2021
16-3-2021
 2817
2817




+
-
20
How Did Interrupted Genes Evolve?
KEY CONCEPTS
-An interesting evolutionary question is whether genes originated with introns or were originally uninterrupted.
-Interrupted genes that correspond either to proteins or to independently functioning noncoding RNAs probably originated in an interrupted form (the “introns early” hypothesis).
-The interruption allowed base order to better satisfy the potential for stem–loop extrusion from duplex DNA, perhaps to facilitate recombination repair of errors.
-A special class of introns is mobile and can insert themselves into genes.
The structure of many eukaryotic genes suggests a concept of the eukaryotic genome as a sea of mostly unique DNA sequences in which exon “islands” separated by intron “shallows” are strung out in individual gene “archipelagoes.” What was the original form of genes?
The “introns early” hypothesis is the proposal that introns have always been an integral part of the gene. Genes originated as interrupted structures, and those now without introns have lost them in the course of evolution.
The “introns late” hypothesis is the proposal that the ancestral protein-coding sequences were uninterrupted and that introns were subsequently inserted into them.
In simple terms, can the difference between eukaryotic and prokaryotic gene organizations be accounted for by the acquisition of introns in the eukaryotes or by the loss of introns in the prokaryotes?
One point in favor of the “introns early” model is that the mosaic structure of genes suggests an ancient combinatorial approach to the construction of genes to encode novel proteins; this is a hypothesis known as exon shuffling. Suppose that an early cell had a number of separate protein-coding sequences; it is likely to have evolved by reshuffling different polypeptide units to construct new proteins. Although we recognize the advantages of this mechanism for gene evolution, that does not necessarily mean that it was the primary reason for the initial evolution of the mosaic structure. Introns might have greatly assisted, but might not have been critical for, the recombination of protein-coding gene segments. Thus, a disproof of the combinatorial hypothesis would neither disprove the “introns early” hypothesis nor support the “introns late” hypothesis.
If a protein-coding unit (now known as an exon) must be a continuous series of codons, every such reshuffling event would require a precise recombination of DNA to place separate proteincoding units in sequence and in the same reading frame (a onethird probability in any one random joining event). However, if this combination does not produce a functional protein, the cell might be damaged because the original sequence of protein-coding units might have been lost.
The cell might survive, though, if some of the experimental recombination occurs in RNA transcripts, leaving the DNA intact. If a translocation event could place two protein-coding units in the same transcription unit, various RNA splicing “experiments” to combine the two proteins into a single polypeptide chain could be explored. If some combinations are not successful, the original protein-coding units remain available for further trials. In addition, this scenario does not require the two protein-coding units to be recombined precisely into a continuous coding sequence. There is evidence supporting this scenario: Different genes have related exons, as if each gene had been assembled by a process of exon shuffling .
FIGURE 1. illustrates the result of a translocation of a random sequence that includes an exon into a gene. In some organisms, exons are very small compared to introns, so it is likely that the exon will insert within an intron and be flanked by functional 5′ and 3′ splice sites. Splice sites are recognized in sequential pairs, so the splicing mechanism should recognize the 5′ splice site of the original intron and the 3′ splice site of the introduced exon, instead of the 3′ splice site of the original intron. Similarly, the 5′ splice site of the new exon and the 3′ splice site of the original intron might be recognized as a pair, so the new exon will remain between the original two exons in the mature RNA transcript. As long as the new exon is in the same reading frame as the original exons (a one-third probability at each end), a new, longer polypeptide will be produced. Exon shuffling events could have been responsible for generating new combinations of exons during evolution.
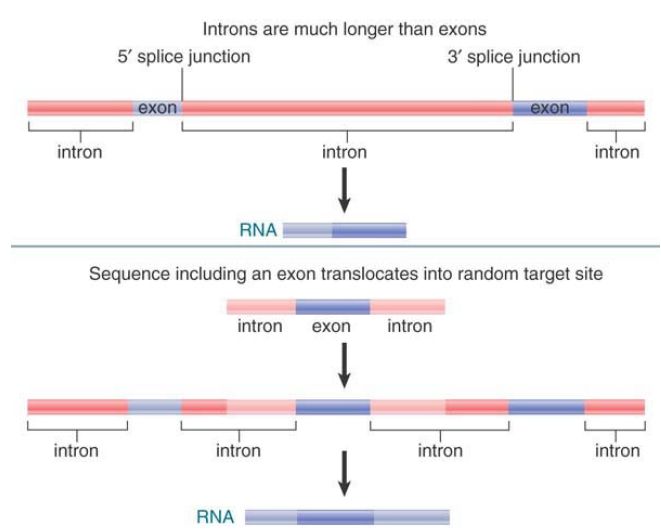
FIGURE 1. An exon surrounded by flanking sequences that is translocated into an intron can be spliced into the RNA product.
Given that it is difficult to envision (1) the assembly of long chains of amino acids by some template-independent process and (2) that such assembled chains would be able to self-replicate, it is widely believed that the most successful early self-replicating molecules were nucleic acids—probably RNA. Indeed, RNA molecules can act both as coding templates and as catalysts (i.e., ribozymes). It was probably by virtue of their catalytic activities that prototypic molecules in the early “RNA world” were able to self-replicate; the templating property would have emerged later.
Many functions mediated by nucleic acid could have competed for genome space in the RNA world. As suggested elsewhere in this text , these functions can be seen as exerting pressures: AG pressure (the pressure for purine-enrichment in exons); GC pressure (the genome-wide pressure for a distinctive balance between the proportions of the two sets of Watson–Crick pairing bases); single-strand parity pressure (the genome-wide pressure for parity between A and T, and between G and C, in single-stranded nucleic acids); and, probably related to the latter, fold pressure (the genome-wide pressure for single-stranded nucleic acid, whether in free form or extruded from duplex forms, to adopt secondary and higher-order stem–loop structures). For present purposes, the functions served by these pressures need not concern us. The fact that the pressures are so widely spread among organisms suggests important roles in the economy of life (survival and reproduction), rather than mere neutrality.
To these pressures competing for genome space would have been added pressures for increased catalytic activities, ribozyme pressure being supplemented or superseded by protein pressure (the pressure to encode a sequence of amino acids with potential enzymatic activity) after a translation system had evolved. Mutation that happened to generate protein-coding potential would have been favored, but would also be competing against preexisting nucleic acid level pressures. In other words, exons might have been latecomers to an evolving molecular system. Given the redundancy of the genetic code, especially at the third base positions of codons, accommodations could have been explored in the course of evolution so that a protein-encoding region would, to a degree, have been subject to selection by nucleic acid pressures within itself. Thus, coding sequences could be selected for both their protein-coding potential and their effects on DNA structure.
Constellations of exons that were slowly evolving under negative selection would have been able to adapt to accommodate nucleic acid pressures. Exon sequences that could accommodate both protein and nucleic acid pressures would have been conserved. However, those evolving more rapidly under positive selection would not have been able to afford this luxury. Thus, some nucleic acid level pressures (e.g., fold pressure) would have been diverted to neighboring introns, resulting in the conservation of the latter.
Some RNA transcripts perform functions by virtue of their secondary and higher-order structures, not by acting as templates for translation. These RNAs, which often interact with proteins, include Xist that is involved in X-chromosome inactivation and the tRNAs and ribosomal RNAs (rRNAs) that facilitate the translation of mRNAs.
Generally, these single-stranded RNAs have the same sequence of bases as one strand (the RNA-synonymous strand) of the corresponding DNA. It is important to note that because these RNAs have structures that serve their distinctive functions (often cytoplasmic), it does not follow that the same structures will serve the (nuclear) functions of the corresponding DNAs equally well. Thus, we should not be surprised that, even though there is no ultimate protein product, RNA genes are interrupted and the transcripts are spliced to generate mature RNA products. Similarly, there are sometimes introns in the 5′ and 3′ untranslated regions of pre-mRNAs that must be spliced out.
Therefore, information for the overtly functional parts of genes can be seen as having had to intrude into genomes that were already adapted to numerous preexisting pressures operating at the nucleic acid level. A reconfiguration of pressures usually could not have occurred if the genic function-encoding parts existed as contiguous sequences. The outcome was that DNA segments corresponding to the genic function-encoding parts were often interrupted by other DNA segments catering to the basic needs of the genome. A further fortuitous outcome would have been a facilitation of the intermixing of functional parts to allow the evolutionary testing of new combinations.
Apart from these pressures on genome space, there are selection pressures acting at the organismal level. For example, birds tend to have shorter introns than mammals, which has led to the controversial hypothesis that there has been selection pressure for compaction of the genome because of the metabolic demands of flight. For many microorganisms (such as bacteria and yeast), evolutionary success can be equated with the ability to rapidly replicate DNA. Smaller genomes can be more rapidly replicated than larger ones, so it might be the pressure for compaction of genomes that led to uninterrupted genes in most microorganisms.
Long protein-encoding sequences had to accommodate numerous genomic pressures in addition to protein pressure. There is evidence that introns have been lost from some members
of gene families.
In the case of the actin gene family, it is sometimes not clear whether the presence of an intron in a member of the family indicates the ancestral state or an insertion event. Overall, current evidence suggests that genes originally had sequences now called introns but can evolve with both the loss and gain of introns.
Organelle genomes show the evolutionary connections between prokaryotes and eukaryotes. There are many general similarities between mitochondria or chloroplasts and certain bacteria because those organelles originated by endosymbiosis, in which a bacterial cell lived within the cytoplasm of a eukaryotic prototype. Although there are similarities to bacterial genetic processes—such as protein and RNA synthesis—some organelle genes possess introns and therefore resemble eukaryotic nuclear genes. Introns are found in several chloroplast genes, including some that are homologous to E. coli genes. This suggests that the endosymbiotic event occurred before introns were lost from the prokaryotic lineage.
Mitochondrial genome comparisons are particularly striking. The genes of yeast and mammalian mitochondria encode virtually identical proteins in spite of a considerable difference in gene organization. Vertebrate mitochondrial genomes are very small and extremely compact, whereas yeast mitochondrial genomes are larger and have some complex interrupted genes. Which is the ancestral form? Yeast mitochondrial introns (and certain other introns) can be mobile—they are independent sequences that can splice out of the RNA and insert DNA copies elsewhere—which suggests that they might have arisen by insertions into the genome. Even though most evidence supports “introns early,” there is reason to believe that, in addition to the introduction of mobile elements, ongoing accommodations to various extrinsic and intrinsic (genomic) pressures might result, from time to time, in the emergence of new introns (“introns late”).
As for the role of introns, it is easy to dismiss intronic characteristics such as an enhanced potential to extrude stem–loop structures as an adaptation to assist accurate splicing. An analogy has been drawn between the transmission of genic messages and the transmission of electronic messages, in which a message sequence is normally interrupted by error-correcting codes.
Although there is no evidence that similar types of code operate in genomes, it is possible that fold pressure arose to aid in the detection and correction of sequence errors by recombination repair. So important would be the latter that in many circumstances fold pressure might trump protein pressure .
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















