























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Cloning
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 5-3-2021
5-3-2021
 3206
3206




+
-
20
Cloning
KEY CONCEPTS
-Cloning a fragment of DNA requires a specially engineered vector.
-Blue/white selection allows the identification of bacteria that contain the vector plasmid and vector plasmids that contain an insert.
Cloning has a simple definition: To clone something is to make an identical copy, whether it is done by a photocopy machine on a piece of paper, cloning Dolly the sheep, or cloning DNA, which is discussed here. Cloning can also be considered an amplification process, in which we currently have one copy and we want many identical copies. Cloning DNA typically involves recombinant DNA.
This also has a simple definition: a DNA molecule from two (or more) different sources.
To clone a fragment of DNA, we must create and copy a recombinant DNA molecule many times. There are two different DNAs needed: a vector, or cloning vehicle, and an insert, or the molecule to be cloned. The two most popular classes of vectors are derived from plasmids and viruses, respectively.
Over the years, vectors have been specifically engineered for safety, selection ability, and high growth rate. “Safety” means that the vector will not integrate into a genome (unless engineered
specifically for that purpose) and the recombinant vector will not autotransfer to another cell. (We discuss selection later.) In general, about a microgram of vector DNA will be ligated with
about a microgram of the insert DNA that we want to clone. Both the vector and insert should be restricted with the same restriction endonuclease to create compatible DNA ends.
Let us now examine the details and the variables that will affect the process, beginning with the insert—the DNA fragment that we want to amplify. The insert could come from one of many different sources, such as restricted genomic DNA—either size selected on an agarose gel or unselected, a larger fragment from another clone to be subcloned (i.e., taking a smaller part of the larger fragment), a PCR fragment , or even a DNA fragment synthesized in vitro. The size and the nature of the fragment ends must be known. Are the ends blunt or do they have overhanging single strands (recall the section “Nucleases” earlier in this chapter), and if so, what are their sequences? The answer to this question comes from how the fragments were created (what restriction enzyme[s] were used to cut the DNA, or what PCR primers were used to amplify the DNA).
The vector is selected based on the answers to these questions. For this exercise, a common type of plasmid cloning vector called a blue/white selection vector is used, as shown in FIGURE 1. This vector has been constructed with a number of important elements.
It has an ori, or origin of replication , to allow plasmid replication, which will provide the actual amplification step, in a bacterial cell. It contains a gene that codes for resistance to the antibiotic ampicillin, ampr , which will allow selection of bacteria that contain the vector. It also contains the E. coli lacZ gene (see the chapter titledThe Operon), which will allow selection of an insert DNA fragment in the vector.
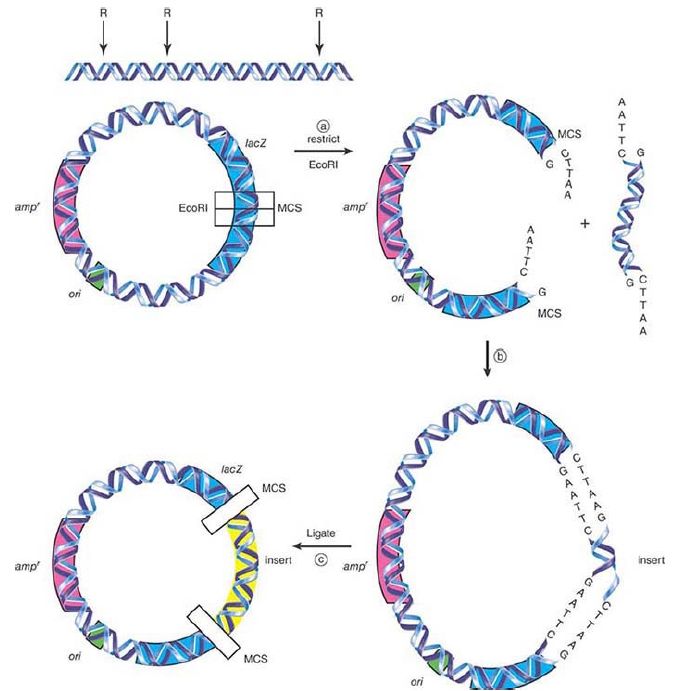
FIGURE 1. (a) A plasmid that contains three key sites (an origin of replication, ori; a gene for ampicillin resistance, ampr ; and lacZ with an MCS), together with the insert DNA to be cloned, is restricted with EcoR1. (b) Restricted insert fragments and vector will be combined and (c) ligated together. The final pool of this DNA will be transformed into E. coli.
The lacZ gene has been engineered to contain a multiple cloning site (MCS). This is an oligonucleotide sequence with a series of different restriction endonuclease recognition sites arranged in tandem in the same reading frame as the lacZ gene itself. This is
the heart of blue/white selection. The lacZ gene codes for the β-galactosidase (β-gal) enzyme, which cleaves the galactoside bond in lactose. It will also cleave the galactoside bond in an artificial substrate called X-gal (5-bromo-4-chloro-3-indolyl-beta-Dgalactopyranoside), which can be added to bacterial growth media and has a blue color when cleaved by the intact enzyme. If a fragment of DNA is cloned (inserted) into the MCS, the lacZ gene will be disrupted, inactivating it, and the resulting β-gal will no longer be able to cleave X-gal, resulting in white bacterial colonies rather than blue colonies. This is the blue/white selection mechanism.
Let us now begin the cloning experiment. Following along in Figure 1., both the vector and the insert are cut with the same restriction enzyme in order to generate compatible single-stranded sticky ends. The variables here are the ability to select different enzymes that recognize different restriction sites as long as they generate the same overhang sequence. An enzyme that makes a blunt cut can also be used, although that will make the next step, ligation, less efficient, but still doable. Two completely different ends with different overhangs can also be used if an exonuclease is used to trim the ends and produce blunt ends. (Continuing with the same reasoning, randomly sheared DNA can also be used if the ends are then blunted for ligation.) If forced to use a type I or type III restriction enzyme, the ends must also be blunted. An important alternative is to use two different restriction enzymes that leave different overhangs on each end. The advantages to this are that neither the vector nor the insert will self-circularize, and the orientation of how the insert goes into the vector can be controlled; this is called directional cloning. Select the vector that has the appropriate restriction endonuclease sites.
The next step is to combine the two pools of DNA fragments, vector and insert, in order to connect or ligate them. A 5- or 10-to-1 molar ratio of insert to vector is usually used. If you use too much vector, vector–vector dimers will be produced. If you use too much insert, multiple inserts per vector will be produced. The size of the insert is important; too large (over ~10 kilobases [kb]) an insert will not be efficiently cloned in a plasmid vector, which will necessitate using an alternative virus-based vector. Ligation is often performed overnight on ice to slow the ligation reaction and generate fewer multimers.
The pool of randomly generated ligated DNA molecules is now used to “transform” E. coli. Transformation is the process by which DNA is introduced into a host cell. E. coli does not normally undergo physiological transformation. As a result, DNA must be forced into the cell. There are two common methods of transformation: washing the bacteria in a high salt wash of calcium chloride (CaCl2 ), or electroporation, in which an electric current is applied. Both methods create small pores or holes in the cell wall.
Even with these methods, only a tiny fraction of bacterial cells will be transformed. The strain of E. coli is important. It should not have a restriction system or a modification system to methylate the incoming DNA. The strain should also be compatible with the blue/white system, which means that it should contain the α-complementing fragment of LacZ (the lacZ gene contained in most plasmids does not function without this fragment). DH5α is a commonly used strain.
Transformation results in a pool of multiple types of bacteria, most of which are not wanted because they either contain a vector with no insert or have not taken up any DNA at all. Select the handful of bacteria that contain recombinant plasmids from the millions that do not. The transformed bacterial cells are plated on an agar plate containing both the antibiotic ampicillin and an artificial β-gal inducer called isopropylthiogalactoside (IPTG). The ampicillin in the plate will kill the vast majority of bacterial cells, namely all of those that have not been transformed with the amp plasmid. The remaining bacteria can now grow and form visible colonies. As shown in FIGURE 2., there are two different types of colonies: blue ones that contain a vector without an insert—because β-gal cleaved Xgal into a blue compound—and white ones, for which the inactivated β-gal did not cleave X-gal and so remained colorless.
FIGURE 2. After transformation into E. coli of restricted and ligated vector plus insert DNA, the bacterial cells are plated onto agar plates containing ampicillin, IPTG, and the color indicator, Xgal. Overnight incubation at 37°C will yield both blue and white colonies. The white colonies will be used to prepare DNA for further analysis.
This is not quite the end of the story. False-positive clones, such as those that were formed as vector-only dimers, must be identified and removed. To do so, plasmid DNA must be at least partly purified from each candidate colony, restricted, and run on a gel to check for the insert size. Sequencing the fragment to be absolutely certain a random contaminant has not been cloned is also suggested .
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















