























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Commitment of Pre-mRNA to the Splicing Pathway
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 12-5-2021
12-5-2021
 2512
2512




+
-
20
Commitment of Pre-mRNA to the Splicing Pathway
KEY CONCEPTS
- U1 snRNP initiates splicing by binding to the 5′ splice site by means of an RNA–RNA pairing reaction.
- The commitment complex contains U1 snRNP bound at the 5′ splice site and the protein U2AF bound to a pyrimidine tract between the branch site and the 3′ splice site.
- In cells of multicellular eukaryotes, SR proteins play an essential role in initiating the formation of the commitment complex.
- Pairing splice sites can be accomplished by intron definition or exon definition.
Recognition of the consensus splicing signals involves both RNAs and proteins. Certain snRNAs have sequences that are complementary to the mRNA consensus sequences or to one
another, and base pairing between snRNA and pre-mRNA, orbetwe en snRNAs, plays an important role in splicing.
Binding of U1 snRNP to the 5′ splice site is the first step in splicing. The human U1 snRNP contains the core Sm proteins, three U1- specific proteins (U1-70k, U1A, and U1C), and U1 snRNA. The secondary structure of the U1 snRNA is shown in FIGURE 1. It contains several domains. The Sm-binding site is required for interaction with the common snRNP proteins. Domains identified by the individual stem-loop structures provide binding sites for proteins that are unique to U1 snRNP. U1 snRNA interacts with the 5′ splice site by base pairing between its single-stranded 5′ terminus and a stretch of four to six bases of the 5′ splice site.
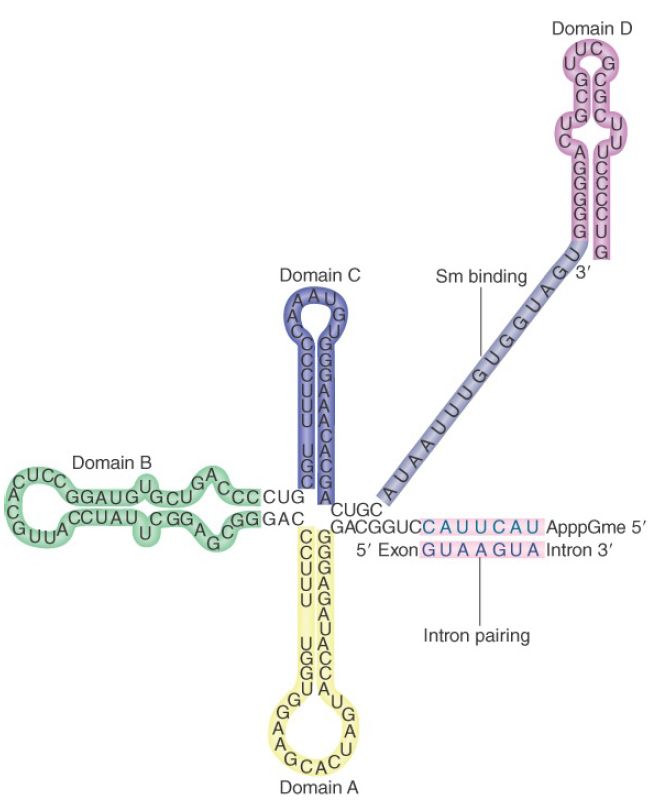
FIGURE 1. U1 snRNA has a base-paired structure that creates several domains. The 5′ end remains single stranded and can base pair with the 5′ splice site.
Mutations in the 5′ splice site and U1 snRNA can be used to test directly whether pairing between them is necessary. The results of such an experiment are illustrated in FIGURE 2. The wild-type sequence of the splice site of the 12S adenovirus pre-mRNA pairs at five out of six positions with U1 snRNA. A mutant in the 12S RNA that cannot be spliced has two sequence changes; the GG residues at positions 5 to 6 in the intron are changed to AU. When a mutation is introduced into U1 snRNA that restores pairing at position 5, normal splicing is regained. Other cases, in which corresponding mutations are made in U1 snRNA to see whether they can suppress the mutation in the splice site, suggest this general rule: Complementarity between U1 snRNA and the 5′ splice site is necessary for splicing, but the efficiency of splicing is not determined solely by the number of base pairs that can form.
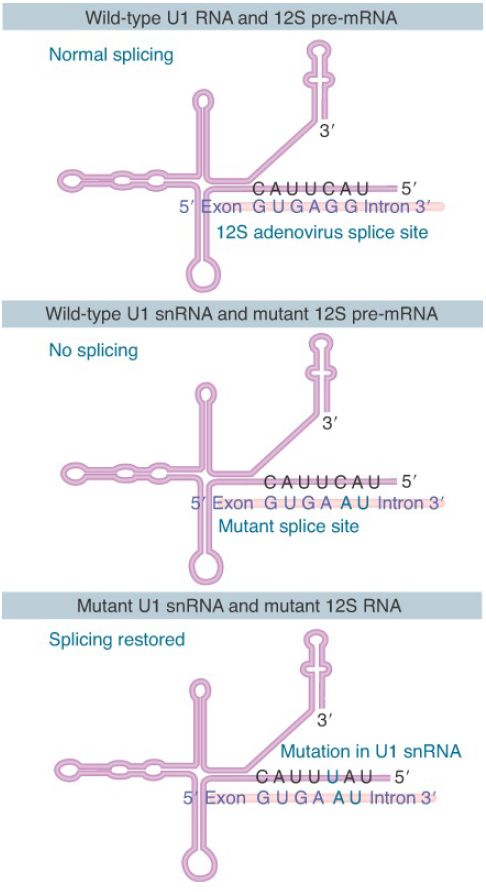
FIGURE 2. Mutations that abolish function of the 5′ splice site can be suppressed by compensating mutations in U1 snRNA that restore base pairing.
The U1 snRNA pairing reaction with the 5′ splicing is stabilized by protein factors. Two such factors play a particular role: The branch point binding protein (BBP, also known as SF1) interacts with the branch point sequence, and U2AF (a heterodimer consisting of U2AF65 and U2AF35 in multicellular eukaryotic cells or Mud2 in the yeast S. cerevisiae) binds to the polypyrimidine tract between the branch point sequence and the invariant AG dinucleotide at the end of each intron. Each of these binding events is not very strong, buttogethe r they bind in a cooperative fashion, resulting in the formation of a relatively stable complex called the commitment complex.
The commitment complex is also known as the E complex (E for “early”) in mammalian cells, the formation of which does not require ATP (compared to all late ATP-dependent steps in the assembly of the spliceosome; see the section later in this chapter titled The Spliceosome Assembly Pathway). Unlike in yeast, however, the consensus sequences at the splice sites in mammalian genes are only loosely conserved, and consequently additional protein factors are needed for the formation of the E complex.
The factor or factors that play a central role in this and other spliceosome assembly processes are SR proteins, which constitute a family of splicing factors that contain one or two RNArecognition motifs at the N-terminus and a signature domain rich with multiple Arg/Ser dipeptide repeats (called the RS domain) at their C-terminus. Their RNA-recognition motifs are responsible for sequence-specific binding to RNA, and the RS domain can bind to both RNA and other splicing factors via protein–protein interactions, thereby providing additional “glue” for various parts of the E complex.
As illustrated in FIGURE 3, SR proteins can bind to the 70-kD component of U1 snRNP (the U1 70-kD protein also contains an RS domain, but it is not considered a typical SR protein) to enhance or stabilize its base pairing with the 5′ splice site. SR proteins can also bind to 3′ splice site–bound U2AF (an RS domain is also present in both U2AF65 and U2AF35). These protein–protein interaction networks are thought to be critical for the formation of the E complex. SR proteins copurify with the Pol II complex and are able to kinetically commit RNA to the splicing pathway; thus they likely function as the splicing initiators in multicellular eukaryotic cells.
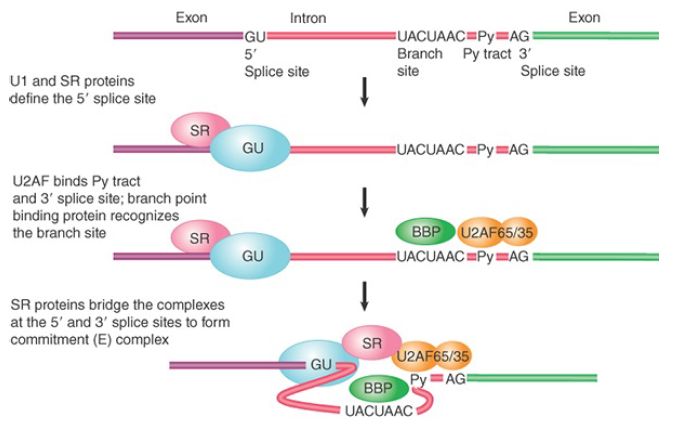
FIGURE 3. The commitment (E) complex forms by the successive addition of U1 snRNP to the 5′ splice site, U2AF to the pyrimidine tract/3′ splice site, and the bridging protein SF1/BBP.
Typical SR proteins are neither encoded in the genome of S.cerevisiae nor needed for splicing by the organism where the splicing signals are nearly invariant, but they are absolutely essential for splicing in all multicellular eukaryotes where the splicing signals are highly divergent. The evolution of SR proteins in multicellular eukaryotes likely contributes to high-efficacy and highfidelity splicing on loosely conserved splice sites. The recognition of functional splice sites during the formation of the E complex can take two routes, as illustrated in FIGURE 4. In S. cerevisiae, where nearly all intron-containing genes are interrupted by a single small intron (between 100 and 300 nucleotides in length), the 5′ and 3′ splice sites are simultaneously recognized by U1 snRNP, BBP, and Mud2, as discussed earlier. This process is referred to as intron definition and is illustrated on the left of Figure 4.
(Note that the intron definition mechanism applies to small introns in multicellular eukaryotic cells, and thus the figure is drawn with the nomenclature for mammalian splicing factors involved in the process.)
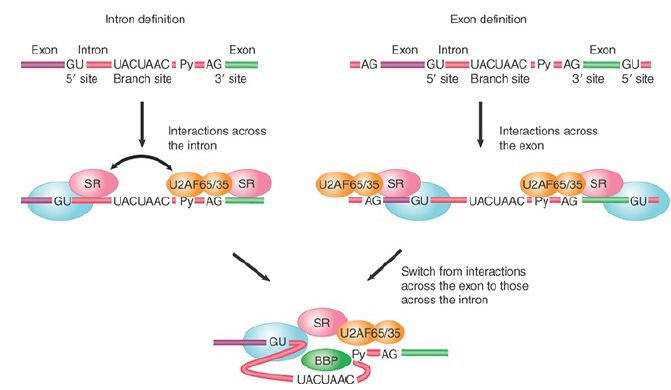
FIGURE 4. The two routes for initial recognition of 5′ and 3′ splice sites are intron definition and exon definition.
In comparison, introns are long and highly variable in length in multicellular eukaryotic genomes, and there are many sequences that resemble real splice sites in them. This makes the paired recognition of the 5′ and 3′ splice sites inefficient, if not impossible.
The solution to this problem is the process of exon definition, which takes advantage of normally small exons (between 100 and 300 nucleotides in length) in multicellular eukaryotic cells. As shown on the right side of Figure 4, during exon definition the U2AF heterodimer binds to the 3′ splice site and U1 snRNP base pairs with the 5′ splice site downstream from the exon sequence. This process may be aided by SR proteins that bind to specific exon sequences between the 3′ and downstream 5′ splice sites. By an as yet unknown mechanism, the complexes formed across the exon are then switched to the complexes that link the 3′ splice site to the upstream 5′ splice site and the downstream 5′ splice site to the next downstream 3′ splice sites across introns.
This establishes the “permissive” configuration that allows later spliceosome assembly steps to occur. Blockage of this transition is actually a means to regulate the selection of certain exons during regulated splicing . Finally, the exon definition mechanism mediated by SR proteins also provides a mechanism to only allow adjacent 5′ and 3′ splice sites to be paired and linked by splicing.
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















