























النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الميكروبية
مواضيع عامة في المضادات الميكروبية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

The 5′ End of Eukaryotic mRNA Is Capped
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 9-5-2021
9-5-2021
 2557
2557




+
-
20
The 5′ End of Eukaryotic mRNA Is Capped
KEY CONCEPTS
- A 5′ cap is formed by adding a G to the terminal base of the transcript via a 5′–5′ link.
- The capping process takes place during transcription and may be important for release from pausing of transcription.
- The 5′ cap of most mRNA is monomethylated, but some small noncoding RNAs are trimethylated.
- The cap structure is recognized by protein factors to influence mRNA stability, splicing, export, and translation.
Transcription starts with a nucleoside triphosphate (usually a purine, A or G). The first nucleotide retains its 5′-triphosphate group and makes the usual phosphodiester bond from its 3′ position to the 5′ position of the next nucleotide. The initial sequence of the transcript can be represented as:
5′pppA /G pNpNpNp …
However, when the mature mRNA is treated in vitro with enzymes that should degrade it into individual nucleotides, the 5′ end does not give rise to the expected nucleoside triphosphate. Instead it contains two nucleotides that are connected by a 5′–5′ triphosphate linkage and also bear a methyl group. The terminal base is always a guanine that is added to the original RNA molecule after transcription.
Addition of the 5′ terminal G is catalyzed by a nuclear enzyme, guanylyl-transferase (GT). In mammals, GT has two enzymatic activities, one functioning as the triphosphatase to remove the two phosphates in GTP and the other as the guanylyl-transferase to fuse the guanine to the original 5′-triphosphate terminus of the RNA. In yeast, these two activities are carried out by two separate enzymes. The new G residue added to the end of the RNA is in the reverse orientation from all the other nucleotides:
5′Gppp + 5′pppApNpNp … → Gppp5′–5′ApNpNp … + pp + p
This structure is called a cap. It is a substrate for several methylation events. FIGURE 1 shows the full structure of a cap after all possible methyl groups have been added. The most important event is the addition of a single methyl group at the 7 position of the terminal guanine, which is carried out by guanine-7- methyltransferase (MT).
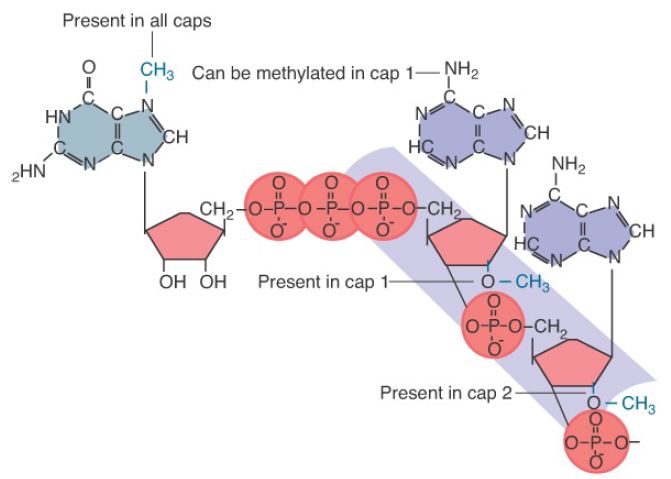
FIGURE 1. The cap blocks the 5′ end of mRNA and can be methylated at several positions.
Although the capping process can be accomplished in vitro using purified enzymes, the reaction normally takes place during transcription. Shortly after transcription initiation, Pol II is paused about 30 nucleotides downstream from the initiation site, waiting for the recruitment of the capping enzymes to add the cap to the 5′ end of nascent RNA. Without this protection, nascent RNA may be vulnerable to attack by 5′–3′ exonucleases, and such trimming may induce the Pol II complex to fall off of the DNA template. Thus, the process of capping is important for Pol II to enter the productive mode of elongation to transcribe the rest of the gene. In this regard, the pausing mechanism for 5′ capping represents a checkpoint for transcription reinitiation from the initial pausing site.
In a population of eukaryotic mRNAs, every molecule contains only one methyl group in the terminal guanine, generally referred to as a monomethylated cap. In contrast, some other small noncoding RNAs, such as those involved in RNA splicing in the spliceosome , are further methylated to contain three methyl groups in the terminal guanine. This structure is called a trimethylated cap.
The enzymes for these additional methyl transfers are present in the cytoplasm. This may ensure that only some specialized RNAs are further modified at their caps. One of the major functions for the formation of a cap is to protect the mRNA from degradation. In fact, enzymatic decapping represents one of the major mechanisms to regulate mRNA turnover in eukaryotic cells . In the nucleus, the cap is recognized and bound by the cap binding CBP20/80 heterodimer. This binding event stimulates splicing of the first intron and, via a direct interaction with the mRNA export machinery (TREX complex), facilitates mRNA export out of the nucleus. Once reaching the cytoplasm, a different set of proteins (eIF4F) binds the cap to initiate translation of the mRNA in the cytoplasm.
 الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
الاكثر قراءة في مواضيع عامة في الاحياء الجزيئي
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة

الآخبار الصحية


مواضيع ذات صلة
 قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة
قسم الشؤون الفكرية يصدر كتاباً يوثق تاريخ السدانة في العتبة العباسية المقدسة "المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة
"المهمة".. إصدار قصصي يوثّق القصص الفائزة في مسابقة فتوى الدفاع المقدسة للقصة القصيرة (نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)
(نوافذ).. إصدار أدبي يوثق القصص الفائزة في مسابقة الإمام العسكري (عليه السلام)



















