
x

هدف البحث
بحث في العناوين
بحث في المحتوى
بحث في اسماء الكتب
بحث في اسماء المؤلفين
اختر القسم
موافق


النبات

مواضيع عامة في علم النبات
الجذور - السيقان - الأوراق
النباتات الوعائية واللاوعائية
البذور (مغطاة البذور - عاريات البذور)
الطحالب
النباتات الطبية
الحيوان
مواضيع عامة في علم الحيوان
علم التشريح
التنوع الإحيائي
البايلوجيا الخلوية
الأحياء المجهرية
البكتيريا
الفطريات
الطفيليات
الفايروسات
علم الأمراض
الاورام
الامراض الوراثية
الامراض المناعية
الامراض المدارية
اضطرابات الدورة الدموية
مواضيع عامة في علم الامراض
الحشرات
التقانة الإحيائية
مواضيع عامة في التقانة الإحيائية
التقنية الحيوية المكروبية
التقنية الحيوية والميكروبات
الفعاليات الحيوية
وراثة الاحياء المجهرية
تصنيف الاحياء المجهرية
الاحياء المجهرية في الطبيعة
أيض الاجهاد
التقنية الحيوية والبيئة
التقنية الحيوية والطب
التقنية الحيوية والزراعة
التقنية الحيوية والصناعة
التقنية الحيوية والطاقة
البحار والطحالب الصغيرة
عزل البروتين
هندسة الجينات
التقنية الحياتية النانوية
مفاهيم التقنية الحيوية النانوية
التراكيب النانوية والمجاهر المستخدمة في رؤيتها
تصنيع وتخليق المواد النانوية
تطبيقات التقنية النانوية والحيوية النانوية
الرقائق والمتحسسات الحيوية
المصفوفات المجهرية وحاسوب الدنا
اللقاحات
البيئة والتلوث
علم الأجنة
اعضاء التكاثر وتشكل الاعراس
الاخصاب
التشطر
العصيبة وتشكل الجسيدات
تشكل اللواحق الجنينية
تكون المعيدة وظهور الطبقات الجنينية
مقدمة لعلم الاجنة
الأحياء الجزيئي
مواضيع عامة في الاحياء الجزيئي
علم وظائف الأعضاء
الغدد
مواضيع عامة في الغدد
الغدد الصم و هرموناتها
الجسم تحت السريري
الغدة النخامية
الغدة الكظرية
الغدة التناسلية
الغدة الدرقية والجار الدرقية
الغدة البنكرياسية
الغدة الصنوبرية
مواضيع عامة في علم وظائف الاعضاء
الخلية الحيوانية
الجهاز العصبي
أعضاء الحس
الجهاز العضلي
السوائل الجسمية
الجهاز الدوري والليمف
الجهاز التنفسي
الجهاز الهضمي
الجهاز البولي
المضادات الحيوية
مواضيع عامة في المضادات الحيوية
مضادات البكتيريا
مضادات الفطريات
مضادات الطفيليات
مضادات الفايروسات
علم الخلية
الوراثة
الأحياء العامة
المناعة
التحليلات المرضية
الكيمياء الحيوية
مواضيع متنوعة أخرى
الانزيمات

Enhancers Contain Bidirectional Elements That Assist Initiation
 المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
المؤلف:
JOCELYN E. KREBS, ELLIOTT S. GOLDSTEIN and STEPHEN T. KILPATRICK
 المصدر:
LEWIN’S GENES XII
المصدر:
LEWIN’S GENES XII
 الجزء والصفحة:
الجزء والصفحة:
 8-5-2021
8-5-2021
 2180
2180




Enhancers Contain Bidirectional Elements That Assist Initiation
KEY CONCEPTS
-An enhancer typically activates the promoter nearest to itself and can be any distance either upstream or downstream of the promoter.
- An upstream activating sequence (UAS) in yeast behaves like an enhancer, but works only upstream of the promoter.
- Enhancers form complexes of activators that interact directly or indirectly with the promoter.
We have largely considered the promoter as an isolated region responsible for binding RNA polymerase. Eukaryotic promoters do not necessarily function alone, though. In most cases, the activity of a promoter is enormously increased by the presence of an enhancer located at a variable distance from the core promoter.
Some enhancers function through long-range interactions of tens of kilobases; others function through short-range interactions and may lie quite close to the core promoter.
One of the first common elements to be described near the promoter was the sequence at −75, now called the CAAT box, named for its consensus sequence. It is often located close to −80,
but it can function at distances that vary considerably from the start point. It functions in either orientation. Susceptibility to mutations suggests that the CAAT box plays a strong role in determining the efficiency of the promoter, but does not influence its specificity. A second common upstream element is the GC box at −90, which contains the sequence GGGCGG. Often, multiple copies are present in the promoter, and they occur in either orientation. The GC box, too, is a relatively common element near the promoter. The concept that the enhancer is distinct from the promoter reflects two characteristics. The position of the enhancer relative to the promoter need not be fixed, but can vary substantially. FIGURE 1 shows that it can be upstream, downstream, or within a gene (typically in introns). In addition, it can function in either orientation (i.e., it can be inverted) relative to the promoter.
Manipulations of DNA show that an enhancer can stimulate any promoter placed in its vicinity, even tens of kilobases away in either direction.
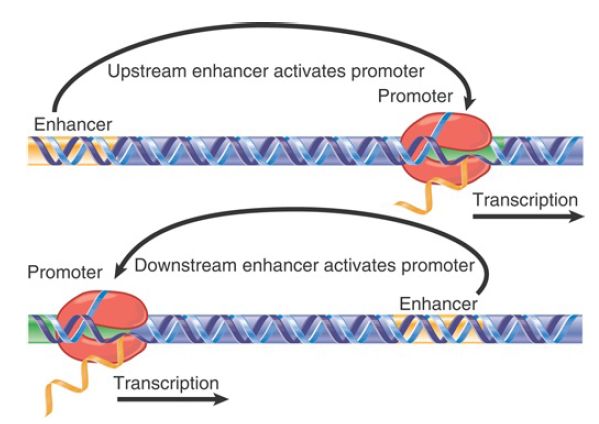
FIGURE 1. An enhancer can activate a promoter from upstream or downstream locations, and its sequence can be inverted relative to the promoter.
Like the promoter, an enhancer (or its alter ego, a silencer) is a modular element constructed of short DNA sequence elements that bind various types of transcription factors. Enhancers can be simple or complex depending on the number of binding elements and the type of transcription factors they bind.
One way to divide up the world of enhancer-binding transcription factors is to consider positive and negative factors. Transcription factors can be positive and stimulate transcription (as activators) or can be negative and repress transcription (as repressors). At any given time in a cell, as determined by its developmental history, that cell will contain a mixture of transcription factors that can bind to an enhancer. If more activators bind than repressors, the element will be an enhancer. If more repressors bind than activators, the element will be a silencer.
Another way to examine the transcription factors that bind enhancers is by function. The first class we will consider is called true activators; that is, they function by both binding specific DNA sites and making contact with the basal machinery at the promoter, either directly by themselves, or, more commonly, through coactivators like Mediator. This class functions equally well on a DNA template or a chromatin template. Two additional classes of activators have completely different mechanisms of activation. One includes activators that function by recruiting chromatin-modification enzymes and chromatin-remodeling complexes. Many activators actually function as true activators and by recruiting chromatin modifiers. The third class includes architectural transcription factors. Their sole function is to change the structure of the DNA, typically to bend it. This can then facilitate bringing together two transcription factors separated by a short distance to synergize. In the next section, Enhancers Work by Increasing the Concentration of Activators Near the Promoter, we examine more closely how the different classes of activators and repressors work together in an enhancer, and in the chapter titled Eukaryotic Transcription Regulation, we examine transcription regulation in more detail. Elements analogous to enhancers, called upstream activating sequences (UASs), are found in yeast. They can function in either orientation at variable distances upstream of the promoter, but cannot function when located downstream. They have a regulatory role: The UAS is bound by the regulatory protein(s) that activates the genes downstream.
Reconstruction experiments in which the enhancer sequence is removed from the DNA and then is inserted elsewhere show that normal transcription can be sustained as long as it is present anywhere on the DNA molecule (as long as no insulators are present in the intervening DNA; see the Chromatin chapter). If a β-globin gene is placed on a DNA molecule that contains an enhancer, its transcription is increased in vivo more than 200-fold, even when the enhancer is several kilobytes upstream or downstream of the start point, in either orientation. It has not yet been discovered at what distance the enhancer fails to work.









 الطاهرة ركن الإسلام الثالث
الطاهرة ركن الإسلام الثالث عناوين أم عنوانات؟
عناوين أم عنوانات؟ أبنائي الطلبة
أبنائي الطلبة EN
EN